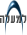למרות שמעלי גירה, כבני אדם וכחד קיבתיים אחרים, משוללים אנזימים צלוליטיים מתוצרת עצמית, הריהם מסוגלים לנצל מזונות מגוונים, ובהם מספוא
עשיר בדופן תא צמחי, המורכב מפחמימות מבניות, צלולוז והמיצלולוז, כמקור לאנרגיה. יכולת זו של מעלי גירה, אשר מאפשרת גם לאדם לנצל מזונות
שאינו יכול לנצלם בעצמו, נובעת מנוכחות מיקרואורגניזמים בכרס מעלי הגירה, שהתמחותם הינה תהליכי פירוק הדופן הצמחי. ממחקרים שנערכו בשנים
האחרונות, עולה כי אוכלוסית הפרוטוזואה והפטריות שבכרס מעלי הגירה אינה שותפה באופן מכריע בתהליכי הפירוק האמורים, אלא קיימים מינים
מסוימים של חיידקים צלוליטיים והמיצלוליטיים, שהתמחו בפירוק הפחמימות המבניות מן הדופן, עובדה המאפשרת את ניצול המזון הצמחי על ידי מעלה
הגירה הפונדקאי.
סידרת מחקרים לאפיון ולזיהוי של החיידקים הדומיננטיים בתהליכי הפירוק, וללימוד מנגנוני הספיחה והאקטיביות , במגוון חומרי מספוא תאיתיים,
נערכה בשנים האחרונות ביחידה המטאבולית שבמכון וולקני. המימצאים פורסמו בכתבי עת בינלאומיים, ולאחרונה בעברית ב"השדה" (כרך ע"ד, חוברת
ז, תשנ"ד) ותמציתם מתפרסמת להלן, יחד עם התיאוריה, שגובשה על פי אותם מימצאים, ביחס לשלבי הפעולה הנחוצים לחיידקי כרס מעלי גירה, לשם
פירוק דופן התא הצימחי.
חמישה זנים עיקריים של חיידקים בודדו ואופינו,במחקר שערכנו, מכרס פרות שניזונו על שחת אספסת. קבוצות מחקר נוספות, מארה"ב, קנדה וסקוטלנד,
מצאו כי אותם חמישה זני חיידקים מפרקים את פחמימות המבנה ברוב חומרי המספוא והליגנוצלולוזות המקובלים בהזנת מעלי גירה. חמשת הזנים
מפורטים להלן:
- : Fibrobacter succinogenes:
חיידק גראם נגאטיבי, בעל תצורת מתג קצר, אשר צלולוז, כסילן, המיצלולוז וכנראה פקטין, הינם הפולימרים העיקריים המשמשים לו סובסטראט.
רוב הקווים הם בעלי כושר פירוק דופן תא במבחנה, כושר ניצול סוכרי הדופן התאי המפורק להתרבות הינו נמוך עד בינוני, ומוצרי התסיסה הינם
אצטאט, סוקצינאט ופורמאט.
- Ruminococcus flavefaciens:
חיידק גראם פוזיטיבי, בעל תצורת כדור, אשר צלולוז, כסילן והמיצלולוז, הינם הפולימרים העיקריים המשמשים לו סובסטראט. לקו 17 כושר גבוה
לפירוק דופן תאי במבחנה (לקווים אחרים כושר נמוך), כושר ניצול סוכרי הדופן התאי המפורק להתרבות הינו בינוני, ומוצרי התסיסה הינם
אצטאט, סוקצינאט, פורמאט ומימן.
- Ruminococcus albus:
חיידק גראם פוזיטיבי, בעל תצורת כדור, אשר צלולוז לא גבישי, כסילן והמיצלולוז, הינם הפולימרים העיקריים המשמשים לו סובסטראט. לקו
3SY כושר גבוה לפירוק דופן תאי במבחנה (לקווים אחרים כושר נמוך), כושר ניצול סוכרי הדופן התאי המפורק להתרבות הוא גבוה מאד, ומוצרי
התסיסה הינם אצטאט, אתנול, מימן ופחמן דו חמצני.
- Butyrivibrio fibrisolvens:
חיידק גראם נגאטיבי, בעל תצורת מתג מאורך, אשר כסילן, המיצלולוז, חלבון, פקטין, עמילן וצלולוז לא גבישי, הינם הפולימרים העיקריים
המשמשים לו סובסטראט. לרוב הקווים כושר נמוך עד בינוני לפירוק דופן תאי במבחנה, כושר ניצול סוכרי הדופן התאי המפורק להתרבות הוא גבוה
מאד, ומוצרי התסיסה הינם אצטאט, בוטיראט, פורמאט וכנראה לקטאט.
- Bacterioides ruminlcola:
חיידק גראם נגאטיבי, בעל תצורת מתג קצר, אשר כסילן, המיצלולוז, חלבון, פקטין ועמילן, הינם הפולימרים העיקריים המשמשים לו סובסטראט.
לרוב הקווים כושר מועט לפירוק דופן תאי במבחנה, כושר ניצול סוכרי הדופן התאי המפורק להתרבות הוא גבוה מאד, ומוצרי התסיסה הינם אצטאט,
בוטיראט, ISO-VFA ואמוניום.
בדיקה מורפולוגית של החיידקים במיקרוסקופ אלקטרוני סורק, בטכניקת הכנה חדשנית (שימוש בפריטין קטיוני) לייצוב פני השטח של החיידקים, העלתה
כי שלושת הזנים הראשונים המנויים לעיל, שגודלו על דופן תאי שחת אספסת כמקור בלעדי לפחמימות, התאפיינו בפני שטח חוץ קפסולאריים מגורגרים
ובעלי בליטות, שכונו "תקביצי פוליצלולוזום" (ראו תמונה), כינוי שמקורו בדמיון המורפולוגי שבין הבליטות שעל פני שטח חיידקי הכרס, לבין
מבנים דומים שכונו כך בחיידק
Clostridium thermocellum ובחיידקים צלוליטיים אחרים.

חיידקי Fibrobacter succinogenes 2BL הספוחים על דופן תאי קש באזור של פיונית טבעית. שים לב לפתיחת תלכידי הצלולוזום
והפיכתם לשלוחות שקושרות את החיידק לדופן.
(הגדלה פי 35,000)
אותם שלושה זני חיידקים, כאשר גודלו, לעומת זאת, על סוכרים מסיסים, צלוביוז או גלוקוז, כמקור פחמימות בלעדי, הפגינו פני שטח חלקים, ללא
תקביצי פוליצלולוזום. כאשר נבדק כושר החיידקים להסתפח לצלולוז, לאחר שהורגלו לפני הספיחה למצע גידול המבוסס על דפנות תאים של אספסת או
למצע שכלל צלוביוז מסיס כמקור פחמימות בלעדי, נתגלה כי החיידקים שהורגלו לגדילה על דופן תא צמחי, היו בעלי כושר ספיחה לצלולוז, גדול מאשר
זה של החיידקים שגודלו על סוכר מסיס. שימוש במיקרוסקופ אלקטרוני סורק, בהמשך המחקר, לימד כי גידול זני חיידקים אלו על מצע מעורב, שכלל
דופן תאי אספסת עם צלוביוז מסיס, גרם לאפיון פני השטח של החיידקים הספוחים לחלקיקי דופן התא, בהופעת תקביצי פוליצלולוזום.
ממימצאים אלה ניתן היה להסיק, כי החיידקים הצלוליטיים הספוחים לדופן התא, עוברים כתוצאה מן המגע בחלקיקי דופן התא, אינדוקציה גנטית
חיובית הגורמת לייצור תקביצי הפוליצלולוזום, תכונה שמועברת לדור החיידקים הבא, ומסייעת בתהליכי הספיחה, ההתישבות על דופן התא ופירוקו.
לעומת זאת, בנוכחות סוכר מסיס, צלוביוז או גלוקוז, אשר מנוצל ביעילות לגדילה, אין החיידקים נזקקים להשקעת האנרגיה הנדרשת לשם ייצור
הפוליצלולוזום, עובדה המותירה את פני השטח החיידקי חלקים ומפחיתה את כושר הספיחה שלהם. מסקנה זו נתמכת כיום בעדויות נוספות שפורסמו
בספרות.
בהמשך המחקר, נסרקו במיקרוסקופ אלקטרוני סורק פני השטח של קווים מסוימים של חיידקי
F. succinogenes וחיידקי
B. fibrisolvens,
שגודלו בתרביות נקיות, ובשילובים של צימדי חיידקים, על מיגוון רחב של דפנות תאים מצימחי מספוא שונים ומליגנוצלולוזות. קווי
F.
succinogenes שגודלו על דפנות תאים מכל המקורות הצמחיים, אופיינו בנוכחות תקביצי הצלולוזום, בעוד שחיידקי
B. fibrisolvens
נתגלו כבעלי פני שטח חלקים, גם כשהיו צמודים לחלקיקים.
הכושר לפרק את דופן התא הצימחי, של חיידקי כרס שונים, נבחן בסידרה של חומרי מספוא וליגנוצלולוזות. קווי
F. succinogenes מצטיינים
ביותר בפירוק דופן התא (בכל המצעים שנבדקו, פורקה הצלולוזה טוב יותר מאשר המיצולולוז), בעוד שקווים של חיידקי
B. fibrisolvens,
R. flavefaciens ו-
R. albus, מפגרים אחריהם.
המסקנות מנסיונות אלה ואחרים, בהם גודלו צימדי זנים של חיידקי כרס על דפנות תאים מבודדות של מיני שחת וליגנוצלולוזות, היו כדלהלן:
בכל שילובי צימדי הזנים שנכדקו, לחיידקי F. succinogenes היה תפקיד מפתח בקביעת דרגת הפירוק של דופן התא.
השפעת גומלין חיובית נמצאה בין זני F. succinogenes לבין זני B. fibrisolvens או B. ruminlcola, ביחס לפירוק
וניצול של פחמימות דופן התא.
השפעת גומלין חיובית נמצאה בין זני R. flavefaciens ו-R. albus לבין זני B. fibrisolvens או
B. ruminlcola, ביחס לפירוק וניצול של פחמימות דופן התא.
השפעת גומלין שלילית נמצאה בין זני F. succinogenes, R. flavefaciens ו-R. albus.
בדיקה במיקרוסקופ אלקטרוני סורק, של אתרי ההתישבות של חיידקי הכרס, בריקמות הגבעול והעלה של חומרי המספוא שנבדקו, לימדה כי מושבות צפופות
של חיידקים קיימות רק בריקמות הפרנכימה, הפלואם וההובלה החשופות, מהן קולפה שיכבת המגן של תאי האפידרמיס, המצופה בקוטין. לעומת זאת,
חיידקים בודדים בלבד נמצאו ברקמות שמוגנות באפידרמיס.
על סמך המימצאים והמסקנות שתוארו, גובשה תיאוריה ביחס למנגנון ולשלבי פעולת החיידקים הצלוליטיים, בעת פירוק חלקיקי דופן תאי צימחי בכרס
של מעלי גירה, והיא מפורטת להלן: < br>
המספוא התאיתי מורטב ומנוטרל ע"י הרוק, ויחדיו עם גריסתו בשיני מעלה הגירה, במהלך תהליכי העלאת הגירה, מוגדל שטח הפנים החשוף של
ריקמות הצמח.
אל חלקיקי הצמח החשופים והשבורים נספחים במהירות חיידקים בכרס מעלה הגירה, תהליך אשר בו משתתפים בעיקר חיידקי F. succinogenes
ו-R. flavefaciens.
אינדוקציה גנטית ליצירת תקביצי פוליצלולוזום חוץ קאפסולריים מתרחשת בחיידקים הצלוליטיים הספוחים לחלקיקים, בעקבות המגע עם דופן התא.
התקביצים המיוצרים בכל החיידקים המתישבים באתר, מסייעים בספיחה לחלקיקים ובפירוק הדופן התאי.
באיזורים חשופים מאפידרמיס מתחילה התרבות מהירה של חיידקי R. flavefaciens והתרבות איטית יותר של חיידקי F.
succinogenes.
דרך הקצוות השבורים או הגמצים הטבעיים נעשית חדירה אקטיבית של זני F. succinogenes וחלק מהזנים של R. flavefaciens, אל
תוך החלל התאי הצמחי.
קשרים גליקוזידיים בסיבוני הצלולוז מפורקים, וכן חל פירוק מצומצם יותר של סוכרי ההמיצלולוז המקושרים לליגנין ולפנולים, תוך ריווח
הדופן המישני, בכיוון השכבות 3S, 2S ו1-S.
חיידקים נוספים, בעיקר R. albus, B. fibrisolvens ו-B. ruminlcola, הגדלים במהירות על פרגמנטים קיצרי שרשרת
שמשתחררים מדופן התא, נכנסים לפעולה.
מתפתחת אוכלוסיה מישנית של חיידקים מנצלי חומצה, לרבות מתאנוגנים, באיזור הדופן המפורק.
תוך 24 שעות מפורקות ריקמות הפארנכימה והשיפה, כמו גם חלק מהאפידרמיס והסקלארנכימה וחלק קטן ביותר מן העצה.
תפקיד הפרוטוזואה בפירוק הדופן אינו ברור. כנראה שפטריות הכרס אינן משתתפות בפירוק הדופן במשך 24 השעות הראשונות לשהות חלקיקי הדופן
התאי בכרס.
עבודת מחקר זו נועדה לסייע בהבנת מנגנון פעולת החיידקים הפעילים בפירוק דופן תאי צמחי בכרס מעלי גירה. לעבודה עשויות להיות השלכות חשובות
בכיוונים שונים: היא עשויה לקדם פיתוח צמחי מספוא עתידיים, בעלי ערך תזונתי רב, שיתרום להגדלת תנובת החלב, השומן והחלבון בפרות חלב,
ולגדילה מהירה שאינה מלווה בהשמנה של עגלים ושל טלאים. היא עשויה לקדם הנדסה גנטית בחיידקי הכרס, לשם הגדלת נצילות המספוא הגס בכרס,
וקבלת מוצרי תסיסה בהתאם לצורכי המגדל.