שתיים מן השאלות היסודיות ששואלים אנשי מדעי החיים הן: כיצד נוצרו החיים (והיכן)? וכיצד התפתחו החיים לאחר ההיווצרות הראשונית? לשאלה הראשונה אין לנו עד היום תשובה מלאה, אם כי מקובל להניח שלאחר תהליך של אבולוציה כימית נוצר תא פשוט - יחסית לתאים המוכרים היום - וממנו התפתחו שלוש שושלות התאים המוכרות לנו היום - החיידקים, הארכאונים, והאיקריוטים (כשהאחרונים כוללים את בעלי החיים, הצמחים והאצות, הפטריות והפרוטוזואונים). את השאלה השנייה מסבירה היום בפרוטרוט האבולוציה.
באמצע המאה ה-19, צ'רלס דרווין (Darwin) הוא שהעלה על הכתב בספרו "מוצא המינים" (On the origin of species) את הסברה שכל היצורים נוצרו אלה מאלה בתהליכים של שינויים אקראיים במידע העובר בהורשה (אז כמובן עדיין לא היה ידוע כיצד) וברירה של פרטים המתאימים יותר לסביבתם בעקבות הבדלים בהעמדת צאצאים בין פרטים המותאמים טוב יותר לסביבתם לבין אלה המותאמים פחות.
כיום כבר מוכר החומר התורשתי (DNA או RNA) וגם מוכרים השינויים החלים בו - המוטציות, המאפשרות את הופעת השינויים האקראיים הנבררים בלחץ תנאי הסביבה. כמו כן ידוע היום, שמלבד השינויים הגנטיים החלים בפרט מסוים, הוא יכול "לרכוש" מידע גנטי חדש באמצעות קליטת חומר תורשתי משכניו. חיידקים יכולים לקלוט חומר תורשתי כפלסמידים (קטעי DNA המשוכפלים בתא החיידק בנפרד מהכרומוזום שלו) ומגוון רחב של אורגניזמים קולטים חומר תורשתי זר מנגיפים. תופעה זו מכונה העברה אופקית (Horizontal gene transfer).
חוקרים יכולים להדגים במבחנה תהליכים אבולוציוניים פשוטים, כגון הופעת עמידות בפני החומר האנטיביוטי סטרפטומיצין בחיידקים. דוגמה זו נובעת משינוי של נוקלאוטיד יחיד ב-DNA, שבעקבותיו מוחלפת חומצה אמינית בודדת בחלבון S12 בריבוזום החיידקי. השינוי גורם לכך שהחלבון, האחראי על הדיוק בתהליך יצירת החלבונים בחיידק, מאבד את יכולתו לקשור את הסטרפטומיצין (המפריע לתהליך ההגהה) בעוד שפעילותו המקורית אינה משתנה. פה גם ברור שבנוכחות סטרפטומיצין, יש עדיפות ברורה לחיידקים הנושאים את המוטציה, ולכן רק הם ישרדו ויעמידו צאצאים, ובסופו של תהליך עברנו מאוכלוסיית חיידקים רגישים לסטרפטומיצין לאוכלוסייה עמידה.
לגבי תהליכים מורכבים יותר, כמו הופעת אברונים חדשים, או ביצורים עילאיים יותר - איברים, שבהם ברור שמדובר בצירוף של כמה שינויים גנטיים, יש קושי להראות איך נוצרות צורות הביניים, שבניגוד לתוצר הסופי, אינן מקנות יתרון לפרט הנושא אותן. כיום אנו מכירים בחשיבותה של המקריות ההיסטורית (Historical contingency) בתהליך הופעתה של התכונה החדשה, למרות הקושי להמחישה בתנאי מעבדה.
באמצעות המערכת הניסויית המתוארת להלן, שנעשה בה שימוש באוניברסיטת מישיגן, הצליחו החוקרים להראות התפתחות של תכונה מורכבת יחסית בחיידק Escherichia coli , ואת קיומה של מקריות היסטורית ייחודית (גנטית במקרה זה). בחיידקים אלה חלו שינויים גנטיים מסויימים, שאף שלא הקנו לחיידקים הנושאים אותם יתרון על פני מתחריהם, אפשרו את הופעת התכונה החדשה.
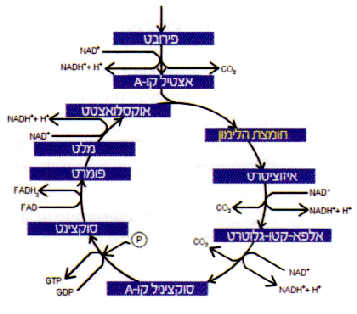
את החיידק E. coli הכרנו במדור זה בגיליון הקודם. אחד המאפיינים הבולטים של חיידקי E. coli יחסית לקוליפורמים האחרים הוא אי-יכולתם של החיידקים לגדול במצע גידול המכיל חומצת לימון כמקור פחמן יחיד. הסיבה לכך אינה מחסור באנזימים המפרקים את חומצת הלימון (הנוצרת באופן טבעי בתוך החיידק במעגל קרבס ובמסלולים אחרים), אלא מחסור בנשא חלבוני, הממוקם במעטפת החיידק, המאפשר את כניסת החומצה לתוך תא החיידק. לכן, בתנאים שגרתיים של מוטגנזה (גרימת שינויים בחומר הגנטי של החיידק), חיפוש מוטנטים Cit+ (בעלי יכולת לגדול על חומצת לימון) של E. coli (שהוא במקורו Cit-) לרוב אינו צולח. אין פלא בדבר, שכן כדי שייווצר מוטנט כזה אין די בשינוי קטן בגן, ומכאן בחלבון קיים, אלא צריך להיווצר גן לנשא חלבוני חדש. במקרים המועטים שבהם נמצאו מוטנטים Cit+ של E. coli, הם נמצאו באוכלוסיות של חיידקים ממינים שונים, והסתבר שהחיידקים המוטנטיים קלטו משכניהם פלסמיד הנושא גן לנשא מַתאים (לאמור - העברה אופקית).
|
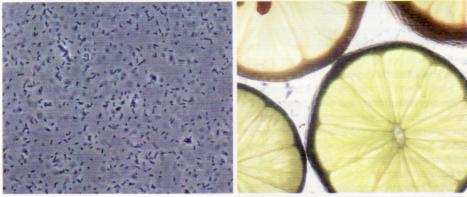
חומצת הלימון
|
|
בשנת 1988 החלו החוקרים זכרי בלאונט (Blount) וכריסטינה בורלנד (Borland), במעבדתו של ריצ'רד לנסקי (Lenski), בניסוי שנתאר להלן. מאז נמשך הניסוי כבר 20 שנה. במהלך הניסוי הממושך הצליחו החוקרים למצוא מוטנט Cit+ של E. coli, באוכלוסייה מבודדת מהסביבה ומחיידקים אחרים - כך שהתפתחות התכונה החדשה נבעה משינויים גנטיים בצאצאי החיידקים המקוריים.
בתחילת הניסוי נזרעו 12 תרביות זהות (שמקורן באותה מושבה) של זן מעבדה של E. coli, שאפשר לזהותו באמצעות סמנים גנטיים ספציפיים.
12 התרביות נזרעו במצע המכיל כמקורות פחמן מעט גלוקוז (25 mg/L) ובעיקר חומצת לימון (סודיום ציטרט - 0.5 gr/L), וגודלו בטלטול בתנאים אווירניים. בכל יום נמהלה התרבית ביחס של 1:100 במצע חדש. הכמות הקטנה של הגלוקוז הגבילה את הגדילה וההתרבות של החיידקים במבחנה, ואיפשרה התרחשות של מוטציות. כשאזל הגלוקוז, הפסיקו החיידקים להתרבות.
כבר בשנים הראשונות של הניסוי חלו בכל תריסר הקווים מוטציות (שונות!), ששיפרו את קצב הגדילה של החיידקים במצע הנתון - עד כ-75% מהר יותר יחסית לחיידקים המקוריים. ואולם, מוטציות המאפשרות כניסה של ציטרט - שאותן אפשר לזהות בקלות, בגלל הגדלה משמעותית של כמות החיידקים במבחנה - לא הופיעו כלל בתריסר התרביות במהלך למעלה מ-30,000 דורות. כל 500 דורות נאספה דגימה מכל תרבית והוקפאה - וכך נוצר מאגר של "חיידקים מאובנים", שהיה אפשר לחזור ולחקור אותם לפי הצורך.
סוף-סוף, לאחר כ-33,000 דורות, התחולל באחת התרביות השינוי המיוחל. באחד החיידקים שבאחד הקווים חלה מוטציה שאפשרה לו לנצל את הציטרט כמקור פחמן. צאצאיו של חיידק זה השתלטו על התרבית בזכות יכולתם לנצל ציטרט. לאחר בדיקת הסמנים הגנטיים המקוריים של החיידקים, כדי לשלול אפשרות של זיהום (כפי שקרה לא מעט במהלך הניסוי, ואז סולקה התרבית המזוהמת והחוקרים חזרו לתרבית קודמת שהוקפאה כדי לשוב ולהמשיך ממנה את הניסוי), ולאחר בדיקה שהחיידקים גדלים גם במצע המכיל ציטרט כמקור פחמן יחיד, יכלו החוקרים להכריז שהמטרה הושגה.
הנחת העבודה של החוקרים היתה, שבחיידק מנצל הציטרט חלה יותר ממוטציה אחת, והשילוב של שתי מוטציות (או יותר) מסוימות אפשר כניסת ציטרט לתא.
כאן הוכנסו מחדש לתמונה החיידקים ה"מאובנים". החוקרים חזרו לתרביות המוקפאות, ובדקו ממתי הפכה לסבירה האפשרות של הופעת מוטציה Cit+. הבדיקה העלתה שב-31,000 הדורות הראשונים של הניסוי לא חל שינוי מקדים כלשהו שיאפשר הופעת מוטנטים Cit+. ואולם, בשלב כלשהו בין דור 31,000 לדור 31,500, חלה מוטציה מקדימה כלשהי באחד החיידקים, שלא השפיעה כלל על הפנוטיפ (קרי, יכולת ניצול הציטרט של החיידקים), אך שכתוצאה ממנה היה אפשר בהמשך לבודד מוטנטים Cit+ בתדירות משתנה. האם גם מוטציה זו, שחשוב עתה לגלות מהי, תלויה במוטציה קודמת (לאור התדירות הנמוכה שבה התרחשה)?
כעת נותר לברר, מהן שתי המוטציות שהתרחשו, וכיצד הן מאפשרות כניסת ציטרט לחיידקים - כך שעדיין רבה המלאכה. המוטציה המקדימה (ואלה המקדימות אותה - שגם אותן יהיה כדאי לנסות ולאתר) המחישה את חשיבות המקריות ההיסטורית בתהליך האבולוציה: ההתפתחות האבולוציונית אינה תוצאה של מוטציות בסדר אקראי. יש חשיבות רבה (שאותה אנו מגלים בדיעבד) לסדר ההתרחשות של המוטציות, או במילים אחרות: למקריות הספציפית של המוטציות בתרבית.
הרבה סבלנות ומשאבים הושקעו בתכנון ובביצוע הניסוי המעניין והארוך הזה, שעודנו נמשך. נקווה שבהמשך נוכל לחזור ולדון בממצאים חדשים ממנו, שירחיבו את הבנתנו את תהליך האבולוציה.
ושאלה נוספת המתעוררת כאן: תקפות ההגדרה והזיהוי של חיידקי ה-E. coli. ברור לנו מדרך הביצוע של הניסוי, שאכן מדובר ב-E. coli. אך אילו היינו מגלים את החיידק הזה בטבע בלא להכיר את תולדותיו, היינו מגדירים אותו כמין, או אפילו כסוג חדש (בגלל היותו מנצל ציטרט). כך גם קרה בעבר עם חיידקי ה-Shigella, הגורמים למחלת הדיזנטריה, שהוגדרו כסוג חדש, בגלל חוסר התנועה האופייני להם (אין להם שוטונים, בניגוד לשאר הזנים של E. coli). רק הבדיקה הגנטית הראתה שגם הם בעצם חיידקי E. coli.
תודה לד"ר שרה שוורץ ולאהוד לם על הערותיהם המועילות.
לקריאה נוספת:
המאמר המקורי:
Zachary D. Blount, Christina Z. Borland, and Richard E. Lenski (2008), Historical contingency and the evolution of a key innovation
in an experimental population of Escherichia coli. PNAS 105 (23) 7899-7906.
קישור לתקציר המאמרהאתר של ריצ'רד לנסקי