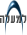נחשפתי לראשונה לקבוצת חיידקים מרתקת זו כסטודנט צעיר למיקרוביולוגיה. החיידקים סקרנו אותי מבחינת מדעית וכן היה בהם להלהיב את הדמיון - היה לנו סרט בו הוצגו נחילי מיקסובקטריה טורפי חיידקים באור אלים במיוחד. אורח החיים שלהם מהווה מודל לתהליכי התפתחות והתמיינות, אולם המיוחד והמעניין הוא התפקיד החשוב של החיים בחברותא, ויחסי הגומלים הבין- תאיים, לא רק במופע הוגטטיבי אלא גם ב"קבלת ההחלטות" הנוגעת לתהליך ההתפתחותי.
החיידק Myxococcus xanthus הינו הנציג הנחקר ביותר בהיבטים הפיסיולוגיים, הגנטיים והמולקולריים של מחזור החיים בקבוצה זו. בקרת ההתפתחות נחקרה בעיקר ע"י Dale Kaiser וקבוצתו (2,1). בחינה גנטית ומולקולתית של מוטנטים רבים, בהם התהליך ההתפתחותי פגום, הראתה שבקרת התהליך והפעלתו דורשים ביטוי של גנים רבים ברצף, כשאחדים מהם זוהו כבקרים של התהליך. עיקרי הממצאים, הדנים בבקרת ההתפתחות בחיידק ע"י צפיפות והרעבה, מוצגים בסקירה שפורסמה לאחרונה ע"י Heidi Kaplan ו-Lynda Plamann (מקור 2) שהחלו במחקר במעבדתו של קייזר והמשיכו בכך במעבדותיהן שלהן. התוצאות, אותן אפרט להלן, מתכנסות למודל לוגי המכליל את התגובה לשני הגורמים הללו.

מחקרים פיסיולוגיים הראו שה"החלטה" על התפתחות מותנית בצירוף של דלדול קרקע המזון וצפיפות מוגדרת של התאים בנחיל. קיום רק אחד מתנאים אילו אינו מספיק לתהליך ההתפתחות: במצב של הרעבה וצפיפות נמוכה, ימשיכו התאים בגידול איטי עד שצפיפותם תגיע לרמת הסף הדרושה להתפתחות. הקריטיות של הצפיפות נובעת מדפוס התזונה של החיידק: התאים הוגטטיביים ניזונים מעיכול חלבונים ע"י פרוטיאזות חוץ-תאיות, שריכוזן במצע הגידול ביחס ישר לצפיפות החיידקים המפרישים אותן. השמרות הנבגים בגופי פרי מוודאת שעם הנביטה יווצרו אוכלוסיות צפופות במידה שתאפשר תזונה יעילה (1).

המוטנטים הפגומים בבקרת ההתפתחות ע"י יחסי גומלין בין תאים,שלא יצרו גופי פרי אלא בנוכחות חיידקים נורמליים או נושאי מוטציות בגנים אחרים, נחלקו לחמש קבוצות: asg, bsg, csg, dsg ו-esg. באמצעות המוטנטים מקבוצה asg, שלא יצרו גופי פרי אלא בנוכחות חיידקים נורמליים ומוטנטים כנ"ל או בהוספת נוזל תרבית "משומש" מחיידקים שייצרו גופי פרי, הוכח קיומו של פקטור חוץ- תאי, שכונה A-signal. הפקטור זוהה כמורכב מפרוטיאזות, המופרשות ע"י החיידקים, וחומצות אמינו, שמציאות כ"א מהן בריכוז מינימום של 0.01mM הכרחית להתפתחות. החוקרים הציעו שהפקטור, המופרש ע"י החיידקים בהרעבה, מהווה "סמן" בין-תאי לצפיפות תאים מתאימה ליצירת גופי פרי.
המוטנטים מקבוצת asg נחלקו לשלושה סוגים - asgA, asgB, asgC. הגנים המתאימים שובטו ואופיינו כאשר asgA זוהה על פי רצף הבסיסים שלו כגן לחלבון המעביר אותות במערכת תרגום אותות מטיפוס two-component system, דגם תרגום והעברת אותות נפוץ בעולם החיידקים. הגן asgB זוהה כגן לחלבון הנקשר ל-DNA ומשתתף בבקרת שיעתוק של גנים. הגן asgC זוהה כגן המקודד פקטור SIGMA, אף הוא חלבון המשתתף בשיעתוק. מבין הסמנים לביטוי גנים במהלך ההתפתחות, נבחר כמודל סמן אחד, 4521, המתבטא תוך 1-2 שעות מתחילת ההתפתחות בתאים נורמליים. ביטוי סמן זה, (שנוצר ע"י איחוי עם גן ל-ביתא גלקטוסידאז שהוחדר ע"י הטרנספוזון Tn5-lac), מותנה בקיום הרעבה ו-A-signal.
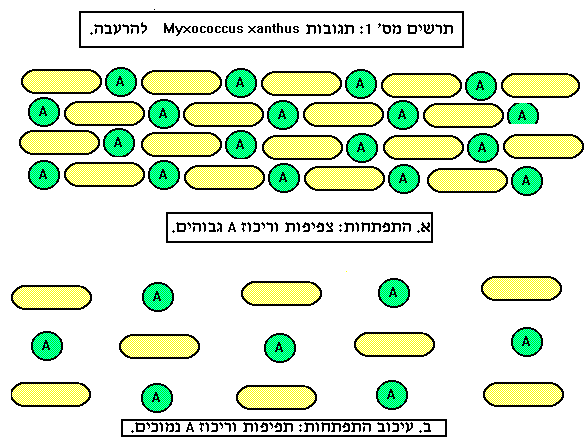
ע"י מוטנטים בהם נעקפת הדרישה לפקטור A בבקרת ההתפתחות בחיידקים נורמליים ומוטנטים הדורשים אותו, זוהו שני גנים שבמצב נורמלי מקודדים דכאנים של תהליך ההתפתחות. גנים אילו כונו sasB ו-sasA. הגן 4521, אותו הזכרנו קודם, מופעל גם כאשר מצטבר בתאים הנוקליאוטיד ppGpp הן בעקבות הרעבה לחומצות אמינו או באופן מלאכותי בגזעים בהם נעשה הטיפול הגנטי המתאים.
נוקליאוטיד זה מופיע בתאי חיידקים במצב של הרעבה לחומצות אמינו, ומזוהה כבקר של סינתיזת RNA בהרעבה ( stringent response) וכנראה גם של תגובות נוספות להרעבה. יש לציין שהגן המייצג 4521 מבוקר ע"י ppGpp באופן ישיר, בנוסף לבקרה ע"י A (מקורות 3,2).
התופעות והממצאים שתוארו נכללים במודל הבא:
במצב של גידול רגיל, הגנים הקשורים לתהליך ההתפתחות מדוכאים במנגנון בו מעורבים תוצרי הגנים sas. עם הידלדלות מצע הגידול, הנוקליאוטיד ppGpp משרה סדרת תגובות להרעבה, ביניהן יצור והפרשת הפרוטיאזות של סמן A (בתווך הגנים ממשפחת asg), המפרקות חלבונים של החיידק ומייצרות את חומצות האמינו שריכוזן מהווה מדד לצפיפות. חומצות האמינו בריכוז מתאים, מנטרלות את הפעולה המדכאה של תוצרי הגנים ממשפחת sas ומאפשרות לתגובה ההתפתחותית להרעבה "לרוץ".
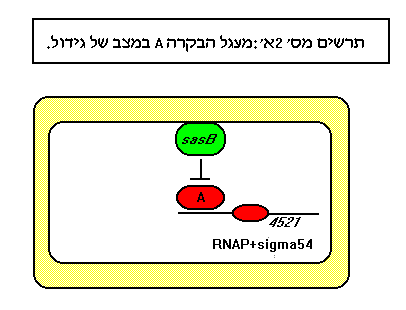
המנגנון המוצע לבקרת יצורו של הסמן A ע"י תוצרי הגנים מקבוצת asg הינו פוספורילציה עצמית של החלבון AsgA המהווה סנסור להרעבה, ומעביר את האות המולקולרי לפקטורי שיעתוק (AsgB ואחרים) הגורמים להפעלת הגנים הדרושים ליצירת הפרוטיאזות והפרשתן.
לסיכום - בקרת הסמן הגנטי 4521 ע"י תגובת ההרעבה מחד וסמן הצפיפות מאידך מראה כיצד החיידק "בוחן" את תנאי הסביבה ומגיב רק כאשר מתקיימים שני התנאים הדרושים. אם נרצה - זהו מנגנון חישובי בו מתבצעת הפעולה הלוגית "AND". כאן ברצוני לציין שהמודל שהוצג כאן אינו שלם ופרטים רבים עודם טעונים בירור. תהליך ההתפתחות החיידק הנדון מורכב ומבוקר ע"י מנגנוני ויסות נוספים לאילו שתוארו כאן.
גנים המבוקרים ע"י צפיפות האוכלוסיה אינה תופעה יחודית למיקסובקטריה. בעולם החיידקים ידועים מקרים נוספים בהם תפקודים הקשורים בסינארגיזם מבוקרים ע"י צפיפות, באמצעות פקטורים המופרשים ע"י החיידקים עצמם, כמו מערכת ההארה בחיידקים מאירים סימביונטים, טוקסינים של Pseudomonas ועוד (4). המיוחד במיקסובקטריה הוא השילוב של צפיפות והרעבה.
השוואה בלתי נמנעת היא ל"עובש" .Dictyostelium disciodeum זהו מיקרואורגניזם איקריוטי, שמחזור חייו, הכולל שלב וגטטיבי בצורת אמבות חד-תאיות, והתקבצות, בעת הרעבה לחומצות אמינו ל- slug רב-תאי שסופו גוף-פרי המכיל נבגים. מחזור החיים דומה מאוד למחזור החיים של M. xanthus. אולם - המנגנונים המולקולתיים הקולטים אותות חיצוניים ומתרגמים אותם לביטוי גנים הם "איקריוטיים" ודומים לאילו שבתאי בעלי חיים (5), בעוד שהמנגנונים המקבילים ב-M. xanthus הם מהדגם החיידקי: לפנינו דגמי פתרון מולקולתיים שונים לאותה בעייה ביולוגית. במלים אחרות - יישום "חישובי" שונה לאותה בעייה לוגית, תוך שימוש במנגנוני תרגום אותות טיפוסיים לכל ממלכה.
רשימת ספרות
- Kim,S.K.,Kaiser,D.,and Kuspa,A, 1992. Control of cell density and intercellular signaling in Myxococcus developmnet. Ann. Rev. Microbiol., 117-139.
- Kaplan,H.B., and L. Plamann, 1996. A Myxococcus xanthus cell density sensing system required for multicellular development. FEMS Microbiol. Letters,139,89-95. (Reprinted in FEMS Microbiology Alert, no. 40).
- Singer,M and Kaiser,D, 1995. Ectopic production of guanosine penta-and tetraphosphate can initiate early developmental gene expression in Myxococcus xanthus. Genes & Dev., 9, 1633-1644.
- Fuqua,W.C.,Winans,S.C.,and Greenberg,E.P., 1994. Quorum sensing in bacteria:the LuxR-LuxI family of cell density-responsive transcriptional regulators. J.Bacteriol., 176, 269 - 275.
- Gross,J.D., 1994. Developmental decisions in Dictyostelium discoideum. Microbiol. Rev., 58, 330-351.