מבוא
במערכות אקולוגיות מדבריות פועל המחסור במים כגורם מעכב לכל הפעילויות הביולוגיות13.
על כן מרוכזת הפעילות במעגלי המינרלים (נוטריינטים) בקרקעות המדבר במשך התקופה
הקצרה שהקרקע מורטבת על-ידי הגשמים22,7. לימוד התהליכים המתרחשים במעגלי המינרלים בזמן הרטבה חשוב מאוד להבנת תפקוד המערכת האקולוגית במדבר. בסוגי קרקע רבים גדלה הכמות הזמינה של פחמן וחנקן עם הרטבת הקרקע26,16, עצמת הנשימה של המיקרואורגניזמים בקרקע בזמן אירועי ההרטבה יכולה להיות גבוהה עד כדי כך שתגרום לירידה בריכוז החמצן12,11. דבר זה מאפשר קיום תהליכים המתרחשים בחוסר חמצן כמו דניטריפיקציה (שחרור חנקן לאטמוספירה בעקבות פעילות מיקרוביאלית, איור 1) גם באדמת המדבר16.
למערכות אקולוגיות רבות במדבר אופיינית כתמיות משמעותית בפיזור הצומח. שני סוגי הכתמים הקיימים בנגב שונים ביניהם בסוג הצומח, דבר המשפיע על כמות החומר האורגני המיוצרת בהם9,3,2:
- כתמי שיחים (מאקרופיטיים) של צמחים עילאיים, המכילים עשבים ושיחים על תלוליות קרקע נמוכות המכוסות בשכבת נשר-צומח21,17,1 (איור 2).
- כתמי קרומי-הקרקע (מיקרופיטיים), שהשכבה העליונה של הקרקע הופכת לקרום קשה ואטום-יחסית למים, והם מצמיחים כמעט רק צמחים ירודים: אצות כחוליות (ציאנובקטריה), אצות ירוקיות, חזזיות וטחבים8,22 (ראה הרחבה בנושא במאמר המבוא ובמאמרו של צעדי בנושא קרומי-הקרקע בחוברת זו).
באזורים בצפון הנגב שלא עברו הפרעות בידי אדם כרעיית יתר, חפירות, ריסוסים בקוטלי-עשבים, עיבודים בכלים חקלאיים ובניית דרכי עפר, אפשר למצוא על גבי השיחים הרבה שבלולים צמודים לענפים המרכזיים, בעיקר מן המין שבלולית מדברית Trochoidea seetzenii, עד כדי עשרות שבלולים לצמח. שבלולים אלה נזונים מחלקים מתים של השיחים ומפטריות וחזזיות הגדלות על גבי הגבעולים והענפים המתים. הפרשות-השבלולים מצטברות מתחת לשיחים ביחד עם החומר האורגני הצמחי. כמות החומר האורגני המיוצרת ליחידת הנוף היא כתמית ומרוכזת בכתמי השיחים, בגלל ההבדלים בין מרכיבי חברת הצומח. מערכת כתמית כזו, שכמות החומר האורגני ממקור צמחי איננה מפוזרת בה באופן שווה שונה ממערכות שהחומר האורגני בהן רצוף (כגון ביערות). שוני זה מוביל למספר שאלות:
- מעגל החנקן במערכת אקולוגית זו, שלא כמו במעגל החנקן הכללי (איור 1), תלוי בשני הכתמים המרכיבים את הנוף המדברי בנגב, ולכל אחד מן הכתמים הללו יש תפקיד שונה במעגל החנקן. מהי ההשתנות של תנועת החנקן בתוך הכתמים וביניהם? ומהו תפקידם של כל אחד משני סוגי הכתמים? האם כתמי קרומי-הקרקע מתפקדים בעיקר כמקור (ספקים) של חנקן, ואילו כתמי השיחים מתפקדים כמיבלע (כצרכנים), הצורך את החנקן?
- מהו מחזור החנקן שבין נשר-צומח, קרקע, בעלי-חיים וצמחים בכתמים השיחיים?
- האם הצטברות של חומר אורגני עשיר בפחמן (מהתאית שבצמחים) ועני בחנקן תוביל לשטף (זרימה) של חנקן מהסביבה אל הנשר כדי לאפשר את פירוקו?
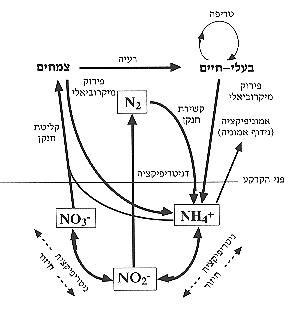
איור 1: סכמת מעגל החנקן בטבע.
מטרות המחקר היו:
- להעריך את הפוטנציאל להוספת חנקן למערכת במסלולים הבאים:
- קשירת חנקן על-ידי מיקרואורגניזמים בקרקע גם בכתמי השיחים וגם בקרומי-הקרקע החשופה;
- תוספת חנקן על-ידי משקעי אבק;
- תוספת חנקן מהפרשות בעלי-חיים;
- תנועת חנקן על-ידי נגר עילי.
- להעריך איבוד חנקן מהכתם השיחי בעקבות פעילות מיקרוביאלית במסלולים הבאים:
- דניטריפיקציה ("אבדן החנקן");
- בריחה של אמוניה;
- ללמוד את תפקוד נשר-הצומח והמיקרואורגניזמים בכתם השיחי והשפעתם על תהליכי מעגל החנקן בקרקעות מדבר.
- לברר את השפעת העונתיות על זרימת החנקן.
שיטות
נאספו קרקע ונשר-אורגני מכתמי השיחים, מתלוליות הקרקע מתחת לשיחים ושכבת
החד-שנתיים המכסים את תלולית הקרקע26. חומר מ-20 תלוליות עורבב ונופה, והחומר שחלקיקיו קטנים מ-0.25 מ"מ נחשב כקרקע. נמדדה הדינמיקה של אמוניום (NH4+) וניטרט (חנקה) (NO3-), דניטריפיקציה וקשירת חנקן ביולוגית ב-30 יום הדגרה במעבדה.
הושוו התגובות להדגרה בין קרקע מקורית לבין קרקע עם תוספת של חומר אורגני מהתלוליות26. התכולה הכללית של פחמן וחנקן והיחס שביניהם נמדדו בקרקע ובנשר-הצומח בתחילת הניסוי. ההדגרה החלה על-ידי הוספת מים מזוקקים מעוקרים כדי להביא את הקרקע לקיבול שדה (כ-22%).
הכלים ובהם הדוגמאות הודגרו בתנאים הדומים לתנאי השדה בחורף בצפון הנגב: 15 שעות חשכה בטמפרטורות של 6-8°C ו-9 שעות הארה בטמפרטורות של 15-18°C. מידת הרטיבות לא השתנתה במשך ההדגרה. הדוגמאות (חד-פעמיות), נבדקו אחרי 2, 10, 20 ו-30 ימים של הדגרה. הדוגמאות טופלו כך שאפשר היה להעריך את השפעת נשר-הצומח על משתני הקרקע. בכל הוצאת דוגמה הורחק החומר האורגני מהחלק העליון של הקרקע ונלקחו דוגמאות הקרקע לבדיקות.
ריכוזי האמוניום והניטרט בקרקע, בנשר-צומח, בהפרשות-השבלולים (מהמין שבלולית מדברית Trochoidea seetzenii) שנאספו מתחת לשיחים, ובנגר העל-קרקעי המגיע לכתם השיחי מיחידת שטח נתונה המכוסה בקרומי-קרקע27 נקבעו על-ידי מיצוי בתמיסת אשלגן-כלורי. לחות הקרקע נקבעה על-ידי שקילה לפני ואחרי ייבוש. ה-pH של הקרקע נקבע בעזרת אלקטרודת זכוכית בתערובת של 1:1 קרקע-מים. דניטריפיקציה נמדדה בכל יום שנלקחה דוגמא24 בעזרת גז כרומוטוגרף.
קשירת החנקן נבדקה ב-4 סוגים של קרומי-קרקע מיקרופיטיים ובמקטע של חומר אורגני מהתלוליות (כתמי השיחים). פוטנציאל קשירת החנקן נקבע על-ידי ריאקציית חיזור אצטילן (C2H2)10. בחישוב החנקן שנקשר השתמשנו ביחס שנמצא לפי הספרות כיחס הנכון לגבי חיזור חנקן אטמוספירי לאמוניום, 3 מולקולות אצטילן למולקולת חנקן אחת.
האפשרות שחנקן, שנקשר על-ידי האצות הכחוליות (ציאנובקטריות) בכתמי קרומי-הקרקע, מועבר אל כתמי השיחים, נבדקה בשלוש צורות: א. על-ידי קשירת החנקן הביולוגית; ב. על-ידי בדיקה כימית של מי הנגר העילי הזורמים מכתמי קרומי-הקרקע אל כתמי השיחים (מחלקות של 0.5 מ"ר של כתמי קרומי-קרקע, שנתחמו על-ידי מחיצות, נאסף הנגר העילי אל מכלי איסוף. המים הועברו לבדיקה כימית של תכולת החנקן); ג. על-ידי מדידת שטף החנקן26.
השוואה נערכה בין נשר-צומח לבין טיפול קרקע כביקורת, ובין זמני הדגרה שונים. הנתונים לגבי קיבוע החנקן נותחו בשתי צורות ניתוח פקטוריאלי של תכנון משתנים, כאשר תקופת ההדגרה והטיפולים היו משתנים בלתי תלויים וייצור האתילן היה משתנה תלוי. השינויים בין הטיפולים וממוצעים של זמני הדגרה נקבעו בעזרת מבחן שיף20.
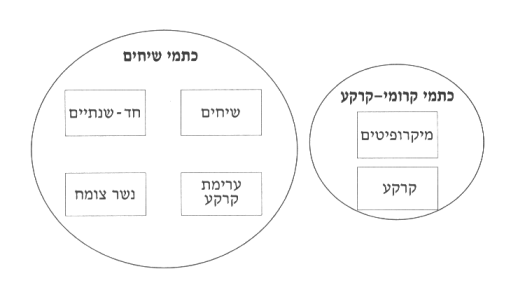
איור 2: סכמה של שני הכתמים המרכיבים את האקוסיסטמה המדברית ומרכיביהם השונים.
תוצאות ודיון
תוספת של חנקן למערכת הכתמית של השיחים יכולה להגיע ולהצטבר ממספר כיוונים:
מקשירת חנקן ביולוגית, ממשקעים יבשתיים (אבק), עם הנגר העילי המגיע מכתמי
קרומי-הקרקע שסביב הכתם השיחי, ומהפרשות-שבלולים. אפשרות קליטה של החנקן
על-ידי קשירת החנקן הביולוגית (קיבוע חנקן אטמוספירי לאמוניום, איור 1), בשני סוגי
הכתמים נבדקה28, ונמצא כי אכן קיימת קשירת חנקן בשני הסוגים. קשירה זו יכולה להתבצע
בכתמי השיחים על-ידי חיידקים קושרי חנקן חופשיים בקרקע, ועל-ידי חיידקים הנמצאים
בסימביוזה עם שורשי צמחי קטניות. בכתמי קרומי-הקרקע מבוצעת רוב קשירת החנקן
על-ידי אצות כחוליות ומיעוטה על-ידי חיידקים קושרי-חנקן חופשיים בקרקע. לא נמצאו
הבדלים משמעותיים בקצב קשירת החנקן בין שני הכתמים. ההבדלים לאחר 12 שעות
(p<0.05) נובעים מתנאי ההארה בניסוי: קושרי החנקן העיקריים בקרומים הם האצות
הכחוליות, ואלה דורשות אור לביצוע הפוטוסינטזה, ורק לאחר שהן מתחילות לבצע
פוטוסינטזה הן מבצעות את תהליך קשירת החנקן28 (איור 3). תרומת החנקן שנמצאה בחישוב
שנתי הייתה 4.32 גרם למ"ר לשנה, תוצאה המתאימה לידוע בנושא זה בספרות22.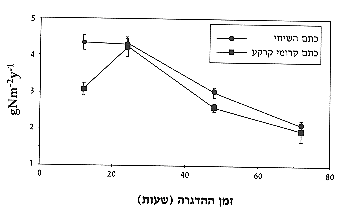
איור 3: קשירת חנקן ביולוגית בכתמי קרומי-הקרקע בהשוואה לכתמי השיחים לאחר 12, 24, 48 ו-72 שעות של הדגרה, ממוצע) שגיאת תקן.
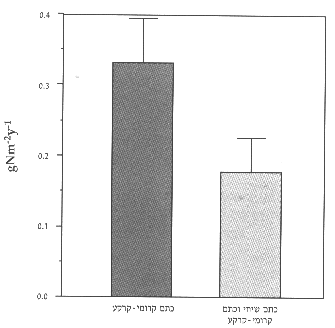
איור 4: כמות החנקן בנגר העילי הדולף ממ"ר של כתמי קרומי-קרקע בהשוואה למ"ר המכיל כתמי קרומי-קרקע וכ-%30 כתם שיחי. ממוצע) שגיאת תקן.
| כמות האבק גרם למ"ר בשנה |
כמות החנקן מיליגרם למ"ר בשנה |
|---|---|
טבלה 1: כמות האבק וכמות החנקן באבק השוקע בצפון הנגב (Offer et al. 1997)
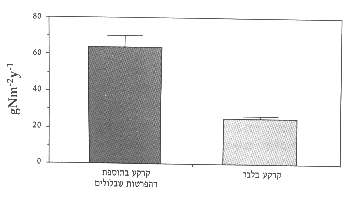
איור 5: כמות החנקן המתקבלת מקרקע בתוספת הפרשות-שבלולים לעומת הקרקע לבדה. ממוצע ) שגיאת תקן.
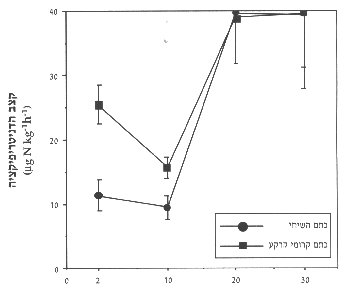
איור 6: קצב הדניטריפיקציה מכתמי שיחים לעומת כתמי קרומי-קרקע לאחר 2, 20 ,10 ו-30 יום הדגרה. ממוצע ) שגיאת תקן.
| זמן (ימים) | pH הקרקע | pH של הקרקע שהודגרה עם נשר-הצומח |
|---|---|---|
טבלה pH :2 של הקרקע שהודגרה וקרקע הביקורת 2, 10, 20 ו-30 יום לאחר ההדגרה
| הטיפול | היחס C:N |
|---|---|
טבלה 3: היחס פחמן לחנקן בנשר-הצומח, בקרקע ובמיקרואורגניזמים בקרקע (בממוצע)
טבלה 4: הביומסה המיקרוביאלית החנקנית בקרקע ובקרקע שהודגרה עם נשר-הצומח 2, 10, 20 ו-30 יום של הדגרה
| ביומסה מיקרוביאלית חנקנית (מיליגרם לק"ג) קרקע |
ביומסה מיקרוביאלית חנקנית (מיליגרם לק"ג) קרקע |
|
|---|---|---|
| זמן (ימים) | קרקע לבדה | קרקע שהודגרה עם נשר-הצומח |
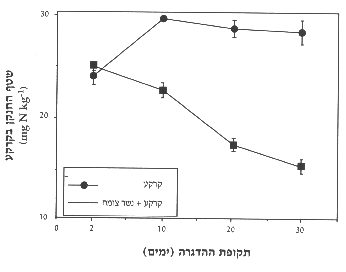
איור 7: שטף החנקן בקרקע לעומת קרקע שהוספה לה שכבת נשר-צומח לאחר 2, 10, 20 ו-30 ימים של הדגרה. ממוצע ) שגיאת תקן.
עונתיות
לפי מעגל החנקן אפשר לחלק את השנה לשלוש עונות בנגב:נשר-צומח מצטבר בזמן העונה היבשה (מאי-אוקטובר). בכתם השיחי מצטבר נשר-הצומח הטרי בקיץ מתחת לשיחים מעשבוניים ומשיירים של השיחים, מהפרשות של שבלולים הנכנסים לפעילות בלילות הטל הרבים בקיץ בנגב29, ומשקיעת משקעי האבק14. בכתם קרומי-הקרקע שוקע אבק, ולעתים, בעת ירידת טל, מאפשרים פני הקרקע הרטובים קשירת חנקן אטמוספירי על-ידי כחוליות קושרות-חנקן (איור 8).
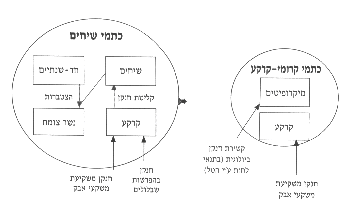
איור 8: תנועת החנקן בשני סוגי הכתמים בתקופה היבשה (קיץ, מאי-אוקטובר).
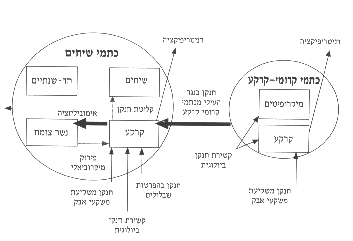
איור 9: תנועת החנקן בשני סוגי הכתמים בתחילת עונת הגשמים (חורף, נובמבר-ינואר).
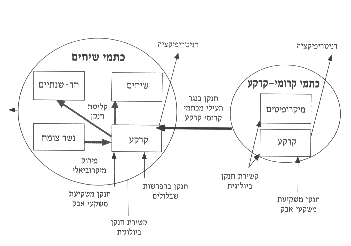
איור 10: תנועת החנקן בשני סוגי הכתמים בתקופת גידול הצומח (אביב, פברואר-אפריל).
סיכום ומסקנות
במערכת האקולוגית המדברית מתרחשים אותם מסלולים במעגל החנקן כמו בכל מערכת
אקולוגית אחרת. אולם במערכת זו נוספו בה מאפיינים הקשורים למבנה הכתמי הטיפוסי
לה, בגלל התפקיד השונה של כל כתם. בעלי-החיים ממלאים תפקיד חשוב במעגל. הנגר העילי
מהווה גורם המקשר בין כתמי קרומי-הקרקע לכתמי השיחים. נשר-הצומח הטרי (המצטבר
בכל קיץ מתחת לשיחים) מהווה מרכיב מרכזי במערכת הכתמית בנוף המדברי. הפוריות
והמגוון הביולוגי בכתמי השיחים נקבעים על-ידי שני גורמים ראשיים: 1. מים (גשר ונגר
עילי); 2. הפירוק המיקרוביאלי של נשר-הצומח ושל הפרשות-השבלולים. נשר-הצומח משמש
גם כמיבלע וגם כמקור לחנקן, והאיזון ביניהם מבוקר על-ידי הפעילות המיקרוביאלית.החשיבות היחסית של המסלולים משתנה לפי עונות השנה (איור 11). נשר-הצומח הטרי הוא קריטי לשימור החנקן בתחילת העונה הרטובה (תחילת עונת הגשמים), והוא מקטין את אבדנו מהמערכת על-ידי נגר עילי, דניטריפיקציה והתאדות אמוניה. החנקן המקובע בעונה זו משוחרר בעונה הבאה כתוצאה מפירוק מיקרוביאלי של נשר-הצומח (כנראה בהשפעת גורמים פיזיקליים כגון עליית הטמפרטורה), ומסייע בכך למערכת בשיא תקופת הצימוח והגידול של הצמחים באביב.
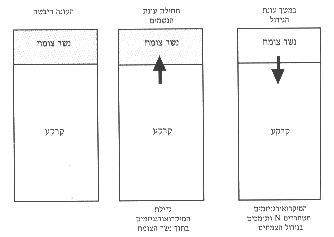
איור 11: סכמת תנועת החנקן בין הקרקע לנשר-הצומח לפי העונות השונות בכתם השיחי.
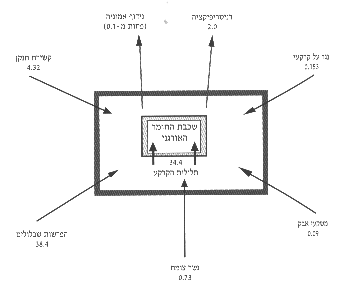
איור 12: סכמת הקליטה והאבדן של החנקן מתלולית הקרקע ושכבת החומר האורגני (נשר-הצומח) בכתמי השיחים (המספרים השונים הם ביחידות של גרם למ"ר לשנה).
תודות
לקרן קיימת לישראל, למר דוד נחמיאס, ד"ר מנחם זקס ומר יצחק משה על תמיכתם במחקר
במסגרת פרוייקט הסווניזציה, לקבוצת המחקר האקולוגית בראשותו של פרופ' משה שחק,
לד"ר ברטרנד בוקן, סול ברנד וסוניה רוזין. כן תודה לד"ר פיטר גרופמן מהמכון לחקר
אקוסיסטמות, מדינת ניו-יורק, על הסיוע בעבודת המעבדה. מאמר זה הנו פרסום של המרכז לחקר המדבור ואקולוגיית השיקום, אוניברסיטת בן-גוריון בנגב, המכון ע"ש יעקב בלאושטיין לחקר המדבר, שדה-בוקר.
ספרות
- Allen, E. B. 1991. Temporal and spatial organization of desert plant communities.
In: Skujins J. (Ed.), Semiarid lands and deserts: soil
resource and reclamation. Marcel Dekker, New York, pp. 193-208.
- Barth, R. C. and Klemmedson, J. O. 1978. Shrub-induced spatial
patterns of dry matter, nitrogen, and organic carbon. Soil Science
society of America J. 42: 804-809.
- Boeken, B. and Shackak, M. 1994. Changes in desert plant
communities in human-made patches and their implications for
management of desertified landscapes. Ecological Application 4, 702-716.
- Danin, A. 1983. Desert vegetation of Israel and Sinai. Cana, Jerusalem.
- Evenari, M. 1985. The desert environment. In: Evenari, M. Noy-Meir, I. And Goodall, D. W. (Eds.).
Hot deserts and arid shrmblands, ecosystems of the world. Elsevier, Amsterdam. Vol. 12A: pp. 1-22.
- Evans, R. D., and Ehleringer, J. R. 1993. A break in the nitrogen
cycle in aridlands? Evidence from 15N of soils. Oecologia 94: 314-317.
- Flie'back, A., Sarig, S. and Steinberger, Y. 1994. Effect of water pulses and climatic conditions
on microbial biomass kinetics and microbial activity in a Yermosol of the center Negev. Arid Soil
Research and Rehabilitation 8: 353-362.
- Friedman, E. I. and Galun, M. 1974. Desert algae, lichens and fungi.
In: Brown, G. W. (Ed.), Desert Biology. Academic Press, London. Vol. 2, pp. 165-212.
- Garcia-Moya, E. and Mekell, C. M. 1970. Contribution of shrubs to
the nitrogen economy of a desert-wash plant community. Ecology 51: 81-88.
- Hardy R. W. F., Burns R. C. and Holsten R. D. 1973. Applications of the acetylene-ethylene assay
for measurement of nitrogen fixation. Soil biology & Biochemistry 5: 47-81.
- Jenkinson, D. S. and Powlson, D. S. 1976. The effects of biocidal
treatments on metabolism in soil-v. Soil Biology and Biochemistry 8: 209-213.
- Kieft, T. L., Soroker, E. and Fierstone, M. K. 1987. Microbial
biomass response to repid increase in water potential when dry soil
is wetted. Soil Biology and Biochemistry 19: 119-126.
- Noy-Meir, I. 1973. Desert ecosystems: environment and producers.
Annual Review of Ecology and Systematic, 4: 25-51.
- Offer, Z., Zaady, E. and Shachak, M. 1998. Aeolian particles input
to microphytic soil crust patches at the northern limit of the Negev
desert. Arid Soil Research and Rehabilitation 12: 55-62.
- Peterjohn, W. T., and Schlesinger, W. H. 1990. Nitrogen loss from
deserts in the southwestern United States. Biogeochemistry 10: 67-79.
- Peterjohn, W. T., and Schlesinger, W. H. 1991. Factors controlling denitrification in a
Chihuahuan desert ecosystem. Soil Science society of America J. 55: 1694-1701.
- Schlesinger, W. H., Reynolds, J. F., Cunningham, G. L., Huenneke, L. F., Jarrell, W. M.,
Virginia, R. A. and Whitford, W. G. 1990.
Biological feedbacks in global desertification. Science 247: 1043-1048.
- Skujins, J. 1981. Nitrogen cycling in arid ecosystems. In: Clark, F. E.
and Rosswall, T. (Eds.), Nitrogen Cycles. Ecological bulltin 33: 477-491.
- Smith J. L., Halvorson J. J. and Bolton H. Jr. 1994. Spatial
relationships of soil microbial biomass and C and N mineralization in
a semi-arid shrub-steppe ecosystem. Soil biology & Biochemistry 26: 1151-1159.
- Sokal, R. R. and Rohlf, F. J., 1981. Biometry, Second Edition. Freeman, San Francisco.
- West, N. E. 1989. Spatial pattern-functional interactions in
shrub-dominated plant communities. In: McKell, C. M. (Ed.), The
Biology and utilization of shrub spp. Academic Press, London. pp. 283-305.
- West, N. E. 19. Structure and function of microphytic soil crusts in
wildland ecosystems of arid to semi-arid regions. Advance Ecological Research 20: 179-223.
- West N. E. and Skujins J., 1977. The nitrogen cycle in North
America cold winter semiarid ecosystems. Oecologia 12: 45-53.
- Yoshinari, T. and Knowles, R. 1976. Acetylene inhibition of nitrous
oxide reduction by denitrificating bacteria. Biophysical Research Communications 69: 705-710.
- Zaady, E., and Shachak, M. 1994. Microphytic soil crust and
ecosystem leakage in the Negev desert. American J. Botany 81: 109.
- Zaady, E., Groffman, P. and Shachak, M. 1996a. Litter as a
regulator of nitrogen and carbon dynamics in macrophytic patches
in Negev desert soils. Soil Biology & Biochemistry 28, 39-46.
- Zaady, E., Groffman, P. and Shachak, M. 1996b. Release and
consumption of snail feces in Negev desert soils. Biology and Fertility of Soil 23: 399-404.
- Zaady, E., Groffman, P. and Shachak, M. 1998. Nitrogen fixation in
macro- and microphytic patches in the Negev desert. Soil Biology and Biochemistry 30: 449-454.
- Zangvill, A. and Druian, P. 1983. Dew in the negev heights. Desert Meteorology papers. Series No. 1.