מי לא נשבה - ולו פעם אחת - בקסמן של בועות סבון: נשפנו קלות בקש שנטבל במי סבון, ואז נוצרה בקצהו בועה, שכלאה בתוכה את הבל פינו החם... והיא הלכה וגדלה; לרגע השתקפו בה פנינו, כמו במראה, לרגע הבהיקה בשלל צבעי הקשת, ואז הופיעו בה כתמים שחורים והיא התפוצצה...
דווקא הכתמים השחורים של בועת הסבון, שבדרך כלל לא שמים לב אליהם, דווקא הם מעניינים אותי עכשיו: באזור הכתמים, עובי קרום הסבון קטן מאורך הגל של האור הנראה, ולכן הם נראים שחורים כשהם מוארים. הם - ולא הבועה הצבעונית - דומים מאוד בעובי לקרומים הדקים שעוטפים את התאים שמהם מורכב גופו של יצור חי. קרומי התאים - דקיקים ושבריריים ככל שיהיו - חושפים כיום בפני החוקרים חלק מחידות החיים.

תמונה 1: בועות סבון
קרום התא - יתרונות הגבול ב"מרק הקדום"
הדעה הרווחת בין חוקרי מקור החיים היא כי אי-שם, ב"מרק הקדום" (המרק הקדום - אגמים וימות שהכילו מלחים ותרכובות אורגניות, שבהן לפי ההשערה, התהוו החיים לראשונה.) לפני כארבעה מיליארדי שנים, הופיעו צברים של מולקולות ענק כתוצאה מתגובות כימיות ספונטניות (שהתרחשו מאליהן). היו אלו - כך משערים - שרשראות של חומצות גרעין (RNA) ושרשראות של חומצות אמינו (חלבונים). שרשראות ה-RNA השתכפלו, ויש שהתגובה בין ה-RNA לחלבונים זירזה את השכפול של מולקולות ה-RNA כך שמספרן גדל במהירות. מסתבר שהחלבונים פעלו כאנזימים (זרזים של תגובות כימיות)!
לפעמים (אולי בתהליכים של ספיחה), הופיע קרום שומני שעטף את צברי המולקולות והבדיל ביניהם לבין צברים אחרים (תמונה 2). סביר להניח כי כאשר שרשראות ה-RNA וה"אנזימים" המשכפלים שלהן היו יחד באותו צבר עטוף. הסיכוי שהן ישוכפלו ביעילות היה גבוה ביחס למקרים שבהם החלבונים נשטפו לים הפתוח. 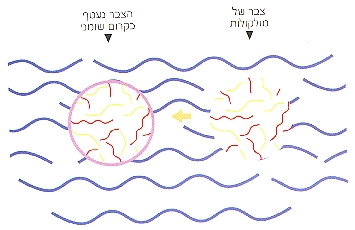
תמונה 2: כך משערים, נוצר האב הקדמון של התא ב"מרק הקדום".
קרום התא - גבול לא מושלם
אין יודעים בפירוט, ואנו יכולים רק לשער, כיצד התרחשה האבולוציה של התא מאביו הקדמון שהיה ב"מרק הקדום". אך גם בתאים המוכרים לנו, הקרום מהווה גבול. הרכב החומרים בתאים החיים כיום, גם ביצורים החד-תאיים וגם ברב-תאיים (כמונו, למשל) שונה לחלוטין מהרכב החומרים מחוץ לתא. למשל, בתוך תאי בעלי-החיים ובאצות החיות בים, קטן ריכוזו של מלח הנתרן פי 20 ויותר מאשר מחוץ לתא. בבעלי-החיים ובצמחים שביבשה גדול ריכוזו של מלח האשלגן בתא פי 50 ויותר מאשר מחוץ לתא, וריכוז הסידן החופשי בתא - בחי ובצומח - קטן פי 10,000 מאשר בחוץ! ועם זאת, התגלה שיונים של נתרן, אשלגן וסידן מתערבבים בין הפנים לבין החוץ, כלומר שהם מסוגלים לחצות את קרום התא. ואף-על-פי-כן, התא החי מצליח להיות בררן. הוא מרכז מולקולות מסוימות ומצליח "להיפטר" ממולקולות אחרות. הקרום הוא שעושה אותו בררן. מלחים חוצים את הקרום השומני במסלולים מיוחדים
הקרום עשוי שכבה כפולה של שומנים (תמונה 3). לא ייפלא אפוא שמולקולות הנמסות בשמן חודרות דרכו בקלות יחסית. אך מי שאי-פעם ערבב שמן ומיץ לימון עם מלח לתיבול הסלט שלו, גילה כבר בוודאי שהמלח אינו נמס בשמן. איך חודרים, אם כן, המלחים דרך קרום התא? התשובה נעוצה במבנה קרום התא. שומני הקרום "ארוזים" בצפיפות בכל אחת משתי שכבותיו: אלו הם הפוספוליפידים, מולקולות בעלי זנבות של חומצות שומן וראשים הידרופילים ("אוהבי מים"). הראשים ההידרופילים הנמסים היטב במים פונים אל הסביבה המימית של פנים התא או של חוץ התא. זנבות חומצות השומן של כל אחת מהשכבות פונים אלו אל אלו ואל פנים השכבה. אך מה שמקנה לקרום את תכונת הבררנות שלו הן מולקולות גדולות של חלבונים ה"תקועות" בין הפוספוליפידים. עוביו של הקרום - כפול בערך מאורך זנבות חומצות השומן - הוא כ-50 אנגסטרום, ואילו חלק מהחלבונים ארוכים מאוד; הם מקופלים, וקיפוליהם מסתלסלים, וחוצים את הקרום לרוחבו מצד לצד הלוך ושוב, כמה פעמים. מספר הקיפולים הללו תלוי בסוג החלבון. חלק מהחלבונים חוצים את הקרום שתי פעמים, ואחרים אף 24! בחלק ממולקולות החלבון הללו, בתוככי קיפוליהן המורכבים, נחבא לו "כיס" או מסלול, שדרכו עוברים יוני המלחים את הקרום השומני בקלות יחסית. כאילו היו בתוך מים, שבהם הם נמסים היטב. לא כל יון מתאים לכל "כיס" או מסלול. יש התאמה לפי מטען היון, חיובי או שלילי - וכך, יש חלבונים המאפשרים מעבר רק לקטיונים (יונים חיוביים) או רק לאניונים (יונים שליליים). ויש חלבונים אחרים הבוררים בין יונים בעלי אותו מטען לפי גודל היון, ומבחינים למשל בין יון אשלגן ובין יון נתרן. 
תמונה 3: קרום התא - פסיפס של חלבונים בתוך שכבה של פוספוליפידים.
בצד שמאל רואים חלבון תעלה פתוח. (איור: גדעון דן)
משאבות: מפלי הריכוזים - סוללה אוגרת אנרגיה
בשם "משאבות" אנו מכנים חלבוני קרום העושים את פעולת הריכוז או הדילול של היונים בתא והיוצרים את ההבדלים בין ריכוזי היונים בפנים התא ומחוצה לו. הבדל זה קרוי מפל ריכוזים. במשאבות מתרחשת תגובה כימית; פירוק של מולקולות ATP, שהן "מטבעות האנרגיה" של התא - וכתוצאה מכך קיפול מסוים בחלבון המשאבה משנה את צורתו. עם שינוי הקיפול מועבר ה"כיס" ובו יון, או יונים אחדים, מצד אחד של הקרום לצד האחר. כך מוציאה משאבת הסידן, למשל, את יוני הסידן מתוך התא החוצה ומקטינה את ריכוזם בתא פי 10,000 ביחס לריכוזם החיצוני. בתאי צמחים וגם בתאי הקיבה שלנו, פעילה משאבה שמוציאה פרוטונים, יוני מימן, ומחמיצה את הסביבה החיצונית של התא.
האנרגיה האצורה ב"מטבעות" ה-ATP מושקעת, אם כן, ביצירת מפלי ריכוזים בין שני צדי קרום התא. לשם מה מנוצלים מפלי הריכוזים?
נשאים: "יבוא אישי"
נשאים הם מולקולות בקרום המזרזות חדירה לתא של חומרים בלתי חדירים. כמו המלח גם הסוכר אינו מסיס בשומני הקרום (זוכרים את הרוטב לסלט?) כיצד אפוא חודר הסוכר לתא? בתאים של בעלי-חיים, בחלבוני קרום מסוימים, קל יותר לקשור מולקולות של סוכר בנוכחות ריכוז גבוה של יוני נתרן. כשהסוכר ויוני הנתרן נקשרים לחלבון-נשא כזה (בצד החיצוני של הקרום, שם ריכוז הנתרן גבוה), בעקבות תנועות קלות של מולקולת החלבון נחשף אתר הקישור של הסוכר והנתרן לסירוגין לכל צד של הקרום, פעם החוצה ופעם פנימה. בצד הפנימי, שם ריכוזו של הנתרן נמוך משתחרר הנתרן, כושר הקשירה של הסוכר לנשא יורד, וגם הסוכר משתחרר (תמונה 4). כך גורם מפל ריכוזי הנתרן ל"יבוא" של מולקולת סוכר אל תוך התא, גם כשריכוז הסוכר בחוץ נמוך מאשר בתא. מולקולות שונות מיובאות לתאים על גבי נשאים. בצמחים, מולקולות קטנות שונות מובאות לתוך התא באופן דומה, תוך ניצול מפל ריכוזי הפרוטונים.

תמונה 4: חלבון נשא בקרום התא (איור: גדעון דן)
תעלות - איזה בזבוז אנרגיה משווע!
דרך מסלולי המעבר בחלבון שמאפשרים מעבר לנתרן - לא "כיסים" אלא "תעלות הנתרן" - יכול יון הנתרן המרוכז מחוץ לתא לדלוף פנימה בלי שתושקע אנרגיה (תמונה 3). כך מתמלא התא ביוני נתרן. הוא הדין בכניסת יוני סידן דרך "תעלות סידן". יוני אשלגן, לעומת זאת, דולפים החוצה דרך "תעלות אשלגן". דרך תעלה יכולים לעבור מיליון ויותר יונים בשנייה, פי 1000 ויותר מכמות היונים העוברים באותו זמן דרך נשא או משאבה. לפנינו פרדוקס: תא חי מייצר ATP ומנצל חלק ניכר ממנו ב"שאיבה" פעילה של יונים ובשמירה על הבדלים בריכוזיהם, ואילו התעלות מאפשרות - וביעילות יתר - דליפה חזרה. איזה בזבוז משווע! האמנם רק בזבוז אנרגיה (ATP) יש בדליפה הזאת דרך תעלות? על כך בהמשך.
למזלנו, גם הבקרה על הדליפות דרך התעלות היא חלק מפעולות החיים שמבצע התא. תגובות כימיות שמשתתפות בהן מולקולות קטנות או יונים, כמו סידן, הנקשרים לחלבוני התעלה - יכולות לקבוע אם התעלות פתוחות או סגורות למעבר של יונים. בחלק מהתעלות גם שדה חשמלי בקרום יכול לקבוע אם הן פתוחות או סגורות. מניין נובע שדה חשמלי בקרום? משאבת הפרוטונים - הסוללה החשמלית של תא הצמח
הקרום השומני של התא פועל כמבודד חשמלי. רק בחלבונים מסוימים יכולים לעבור אלקטרונים - וחלבונים אלו נדירים - יחסית. הנושאים העיקריים של מטען חשמלי דרך הקרום הם היונים העוברים במשאבות, בנשאים ובתעלות. וכך בתאי הצמח, למשל, כאשר משאבת הפרוטונים מוציאה פרוטונים אל מחוץ לתא, מצטבר מטען שלילי בצד הפנימי של קרום התא (במילים אחרות: מצטבר מטען חיובי בצד החיצוני של קרום התא). טעינת הקרום יוצרת בתוכו שדה חשמלי. את הטעינה החשמלית הזאת אפשר למדוד בעזרת שתי אלקטרודות, שאחת מהן, זעירה במיוחד, מחדירים לתא. פעולת משאבת הפרוטונים בצמח יכולה ליצור הפרש מתח חשמלי השווה לחמישית מהמתח החשמלי של סוללת פנס כיס קטנה! ושוב התעלות: פיקוד חשמלי ואיתות חשמלי
מעט מאוד ידוע עד כה על תפקיד ה"סוללה החשמלית" בתאי הצמחים, אבל כבר עתה ברור שה"סוללה" הזאת חיונית ביותר. כאשר השדה החשמלי בקרום התא גדול דיו (שלילי בפנים), נפתחות בו תעלות אשלגן הרגישות לשדה הזה. דרך התעלות נכנסים לתא יוני אשלגן חיוביים המונעים על-ידי המתח החשמלי. כך מצטבר בתא הצמח אשלגן, בניגוד למפל הריכוזים שלו! בלי תהליך זה אין תא הצמח יכול לגדול. תעלות בתמסורת האותות בין גירוי לתגובה
ידוע שבתגובה לגירויים שונים, נרשמים בתאי הצמח איתותים חשמליים. למשל, כשמאירים באור כחול על תת-פסיג של נבט אפונה שצמח בחושך, התארכותו נעצרת בתוך מספר שעות. אבל כמעט מיד, בתוך שניות מתחילת ההארה, נרשמת בתאים שהוארו "דפולריזציה", כלומר ירידה בטעינה החשמלית של הקרום. הסיבה העיקרית לדפולריזציה הזאת היא פתיחה של תעלות בררניות ליוני כלוריד בקרום התאים של התת-פסיג. כיוון שריכוז יוני הכלוריד בתא הצמח תמיד גדול יותר מאשר בחוץ - דולפים יוני הכלוריד החוצה דרך התעלות. כיוון שהם נושאים איתם מטען חשמלי שלילי - נעשית טעינת קרום התא פחות שלילית, או חיובית יותר, בפנים. ידוע שהדפולריזציה הזו חיונית להמשך התגובה: אם חוסמים את תעלות הכלוריד (בחומר כימי מיוחד), התת-פסיג ממשיך להתארך על אף ההארה באור כחול. שלבי הביניים, בין הגירוי לבין עצירת ההתארכות, עדיין לא ידועים. דפולריזציה היא גם שלב ביניים בתהליכים נוספים, למשל בתגובה להורמון הגדילה בצמח, אוקסין, ביצירת סימביוזה בין חיידקים קושרי חנקן לבין שורשים של קטניות, או ביצירת חומרי מגננה בתגובה לפציעה של עלה.
שלבי הביניים בכל התהליכים הללו (תמסורת האותות בין הגירוי לתגובה) הם מהחידות ה"חמות" והמרתקות ביותר של השנים האחרונות בביולוגיה של התא. חלק ניכר מן המחקרים הללו מתמקדים בקרום התא בהיותו הגבול שבין התא, לסביבתו החיצונית. שם, בתוך הקרום או בצמוד לו, קורים השלבים הראשונים של תמסורת האותות.