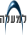הננו-טכנולוגיה התחילה את דרכה בשנות השישים, עם קביעתו של חתן פרס נובל לפיזיקה ריצ'רד פיינמן (Feynman) שלפיה "יש עוד הרבה מקום בתחתית". אנו משתמשים כיום במכונות גדולות, גסות ומגושמות, אשר רובן המכריע מנצל את הדלק המוזרם לתוכן ביעילות נמוכה למדי. מנוע הבערה הפנימית ברכב משפחתי ממוצע, למשל, הופך רק 20% מאנרגיית הדלק המוזרם לתוכו לתנועה; כל השאר מבוזבז על חום ורעש. הננו-טכנולוגיה מציעה לנו אפשרות לעבוד "בתחתית" - ליצור מכונות כה קטנות, שמיליונים מהן יוכלו להימצא על ראש סיכה, ויעילות עד כדי 80% ויותר.
עצם שמה של הננו-טכנולוגיה מייצג את מטרתה: ננו-מטר הוא מיליארדית המטר, כאורכה של מולקולה. טכנולוגיה היא דרכו של האדם ליצור מכונות שיפתרו בעיות עבורו. ייעודה המתקדם ביותר של הננו-טכנולוגיה הוא ייצור מכונות שגודלן כגודל מולקולות - ננו-מטרים אחדים. ננו-מכונות אלו יוכלו לתפעל מולקולות ואטומים יחידים, לחבר ולהפריד ביניהם ובכך להשיג שליטה עילאית בחומרים המיוצרים. יהלום, למשל, עשוי בסך-הכל מאטומי פחמן - אחד היסודות הנפוצים על פני כדור-הארץ. הסיבה לחוזקו העצום היא סידורם המיוחד של האטומים במרחב. אם נוכל לתמרן אטומי פחמן באמצעות ננו-מכונות ולבנות מהם גביש יהלום, הרי שהיהלום יהיה חומר זמין וזול ברגע שמהפכת הננו-טכנולוגיה תגשים את ייעודה. (אמנם, כיום כבר מיוצרים יהלומים מלאכותיים, בתנאים קיצוניים של לחץ וטמפרטורה; אך אלה הם יהלומים קטנים, ולא יריעות או גושים כפי שמקווים ליצור באמצעות הננו-טכנולוגיה.)
אלא שהדרך עוד ארוכה עד למכונות מולקולריות מעשה ידי אדם. עדיין אין ברשותנו כלים שיאפשרו לנו לטפל ביעילות במולקולות ובאטומים בודדים ברמת דיוק כזו, שנוכל להרכיב מהם מכונות כרצוננו. והנה, במקום לעכב את המחקר בתחום עד שהטכנולוגיה שלנו תגיע לרמה כזו של דיוק ויעילות, חוקרים רבים פונים לאפיק מקביל - שימוש במכונות מולקולריות שמקורן אינו בכושר ההמצאה של האדם, אלא בטבע עצמו; מכונות אלו הן האנזימים.
האנזימים הם חלבונים המיוצרים כל העת בתוך כל תא. הם אחראים לפעולות המתרחשות בתא, דוגמת שעתוק המידע הטמון בקוד הגנטי ותרגומו לאנזימים חדשים, או תהליכים מטבוליים סבוכים המאפשרים הפקת אנרגיה ממולקולות פשוטות. אנזימים אלו פועלים במידה רבה על פי עקרונות כימיים שקשה להעביר וליישם בקני-מידה גדולים יותר. יחד עם זאת, ישנם אנזימים אשר פעולתם היא מכנית בעיקרה, ושאפשר לתרגם אותה בפשטות יחסית למושגים המוכרים לנו מעולם ה"מאקרו" - עולם המכונות הגדולות, שאת חוקי הפיזיקה והמכניקה שלו אנו מיטיבים להכיר. אנזימים אלו הם "מנועים מולקולריים", והם עשויים להוות את הדור הראשון של המכונות המולקולריות הנשלטות על-ידי האדם.
שתי קבוצות עיקריות של מנועים מולקולריים הקיימים בטבע הן:
מאז שנות התשעים נערך מחקר אינטנסיבי של המנועים המולקולריים הללו, המתרכז במציאת יישומים הנדסיים לשימוש בהם. בתחום המנועים הסיבוביים המחקר מתמקד בעיקר ב-F-ATPase Synthase. בתחום המנועים הנעים על מסילות, המחקר מתמקד בעיקר בקינזין.
מנועים סיבוביים: F-ATPase Synthase
מקור השם ATPase הוא ביכולתו של האנזים לפרק את מולקולות ה-ATP (אדנוזין טרי-פוספט) ל-ADP (אדנוזין די-פוספט) ולקבוצת פוספט (זרחה) בודדת.
מקור המילה Synthase הוא ביכולתו של אותו אנזים לבצע גם את הפעולה ההפוכה - לחבר אותם מרכיבים למולקולת ATP שלמה. עיקר פעולתו של
האנזים הוא בכיוון יצירת ATP - מולקולות עתירות אנרגיה כימית המשמשות להנעת מכונות מולקולריות אחרות (ראו איור).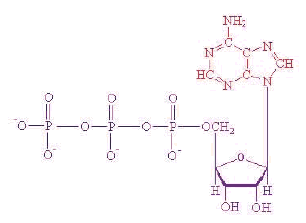
מולקולת ATP, אדנוזין טרי-פוספט, המורכבת מהבסיס החנקני אדנין (בכתום), מהסוכר ריבוז (בוורוד) ומשלוש קבוצות פוספט (זרחה, בכחול). הסרת קבוצת הפוספט הקיצונית היא תהליך משחרר אנרגיה, שתוצריו הם ADP (אדנוזין די-פוספט) וקבוצת פוספט בודדת. ולהפך - קשירת קבוצת פוספט בודדת ל-ADP היא תהליך צורך-אנרגיה שבו נוצר ATP, אדנוזין טרי-פוספט
האנזים F-ATPase Synthase מתבסס על חלק מסתובב - הרוטור - המורכב מ-12 תת-יחידות המרכיבות ביחד גלגל, בדומה ל-12 מדפים של גלגל מים.
כניסת פרוטון למדף אחד גורמת לגלגל להסתובב ב-30 מעלות, ולשחרר פרוטון ממדף אחר. בדרך זו, בכל סיבוב של 30 מעלות, פרוטון אחד נכנס
לגלגל, ופרוטון אחר יוצא ממנו, בתנועה הדומה לזו של גלגל מים. במרכז הגלגל נעוץ ציר מרכזי, ובקצהו מקובע ראש, המוחזק בעזרת ידית כדי
למנוע ממנו להסתובב ביחד עם הציר. תנועת הציר המרכזי בתוך הראש המקובע במקום היא המספקת את האנרגיה הנחוצה לפעולת הראש, אשר מושך
אליו מולקולות ADP וקבוצות פוספט, ומאחד אותן למולקולות ATP. כל סיבוב שלם של 360 מעלות של הגלגל מביא לבניית שלוש מולקולות ATP.
- הרוטור מורכב מ-12 תת-יחידות שביחד יוצרות גלגל, שאותו אפשר לדמות לגלגל בטחנת-מים. הגלגל מקובע בממברנה, ולצדו נמצאת תעלה שדרכה
יכולים לעבור פרוטונים. הפרוטונים נכנסים לכל תת-יחידה בגלגל דרך התעלה, ויוצאים מתוך תת-היחידות בצדה האחר של התעלה. כל כניסה של
פרוטון בצד האחד ושחרור של פרוטון מהצד האחר מביאים לסיבוב של הגלגל ב-30 מעלות.
- אפשר לראות כי בתוך הגלגל מקובע ציר מרכזי, המסתובב גם הוא ביחד עם הגלגל. בקצה האחר של הציר נמצא "ראש" האנזים (בצהוב וסגול). ראש
זה אינו מסתובב, מכיוון שהידית (בצהוב וחום) מחזיקה אותו מן הצד בעמדה קבועה, ואינה מאפשרת לו להסתובב. כאשר הציר המרכזי מסתובב בתוך
הראש, הוא גורם לשינוי תצורה של תת-היחידות המרכיבות את הראש, ובכך מאפשר להן לבצע את פעולתן - לקשור מולקולות ATP ופוספט, ולהפוך
אותן למולקולות ATP - מטבע האנרגיה המניע את החיים.
- ראש האנזים הוא החלק המייצר את ה-ATP, או מפרק אותו. הראש מכיל שלושה אתרים שכל אחד מהם יוצר ATP בפני עצמו, בעקבות תנועת הציר.
בעקבות כל סיבוב שלם של הציר בתוך הראש, נוצרות שלוש מולקולות ATP.
- הציר המרכזי מסתובב בתוך הראש במעברים של 120 מעלות, ובכל סיבוב כזה עובר כל אחד מהאתרים שבראש למצב אחר: 1. מצב פתוח (Open, O) -
האתר קושר מולקולת ADP ומולקולת פוספט, שהן חומרי הגלם ליצירת מולקולת ATP. 2. מצב משוחרר (Loose, L) - האתר מקרב את חומרי הגלם זה
לזה. 3. מצב הדוק (Tight, T) - האתר מפעיל לחץ גדול מספיק כדי לגרום לתגובה בין חומרי הגלם, שתביא ליצירת מולקולת ATP שלמה.
דרך הפעולה של המנוע בתאים מבוססת על מעבר פרוטונים דרך הגלגל כמקור אנרגיה המסובב אותו, והמרת אנרגיה זו ליצירת מולקולות ATP עשירות
באנרגיה. עם זאת, מחוץ לַגוף המנוע יכול לפעול גם בדרך ההפוכה: מולקולות ATP מגיעות לראש, והוא מפרק אותן ובכך גורם לציר המרכזי
להסתובב, ולסובב את הגלגל בכיוון ההפוך. גמישות זו בחומר הדלק של המנוע - מעבר פרוטונים או ATP - היא אחת הסיבות לכך שמנוע מולקולרי
זה הוא כנראה הנחקר ביותר בעולם. ב-1997 פרסם החוקר הירויוקי נוג'י (Noji) דיווח על הצלחתו לצפות בתנועת הסיבוב של המנוע. נוג'י חיבר סיב אקטין פלואורסצנטי לציר המרכזי של המנוע, וצילם את תנועת הסיבוב של הסיב בנוכחות ATP. מחקר זה היה פריצת דרך בתחום המנועים המולקולריים והניע חוקרים רבים להתעניין בנושא.
בעקבות ניסוי זה והעניין הרב שעורר, נוצר ב-2001 השילוב הראשון של מנוע כזה הכולל חלקים אורגניים וסינתטיים. ריקי סונג (Soong) מאוניברסיטת קורנל יצר מדחף ניקל באורך של 1,400 ננו-מטר, הצמיד אותו באמצעות דבק ביולוגי לציר המרכזי של המנוע, והעמיד את המתקן כולו על עמודי סיליקון בגובה 200 ננו-מטר, כדי למנוע מהמדחפים לפגוע בתחתית. מתוך 400 מנועים כאלו שהורכבו, רק חמישה הצליחו לבצע תנועה סיבובית רצופה. עוד רבה, אם כן, הדרך ליצירת מכונות מולקולריות שיפעלו ברמת האמינות הבסיסית שהשגנו במכונות גדולות יותר.
אף שהתאכזב ממספר המנועים הקטן שהצליחו לפעול, הפיק סונג מסקנה חשובה מן הניסוי. מתוך מהירות סיבוב המדחפים וידיעת אורכם, הוא הצליח להעריך את יעילות המנוע המולקולרי ולקבוע את ערכה על כ-80% - פי ארבעה מיעילות מנוע הבערה הפנימית הנמצא ברוב כלי הרכב כיום!
החוקר הייקינג ליו (Liu) השיג ב-2002 התקדמות נוספת בשליטה על המנוע הזעיר, באמצעות התקנה של "מתג הפעלה וכיבוי" במנוע. הייקינג תכנן והוסיף לאזור הראש של המנוע אתרים הקושרים אטומי אבץ (Zn). כאשר האתרים הללו קושרים אליהם אבץ, המבנה המרחבי שלהם משתנה והם משתקים את יכולתו של המנוע להסתובב, בדומה למקל הנתקע בגלגל ומונע את סיבובו. ואכן, בניסויים שנעשו במנוע המולקולרי המשופר, נמצא שהוא הפסיק להסתובב ברגע שהוסף אבץ לתמיסה. המנוע חזר להסתובב כאשר הוסיפו לתמיסה את החומר PNT (פֶנָאנְתרוֹלִין), שמשך אליו את האבץ. כאשר יצא האבץ מן האתרים שבמנוע, הם חזרו לצורתם הרגילה והלא-מפריעה, ואפשרו למנוע להמשיך להסתובב.
מנועים סיבוביים - מתיאוריה למעשה
מנועים מסתובבים מולקולריים יכולים לשמש אותנו בתחומים רבים של הנדסה. כמה משימושיהם האפשריים: - כממירי כוח: מנועים המשתמשים בדלק ביולוגי-כימי (מולקולות ה-ATP) ליצירת כוח סיבובי. אף שכרגע אין לממיר כוח זה שימוש בתעשייה, עם
התקדמות התחום נצפה למצוא יותר ויותר מכשירים העושים שימוש במקורות אנרגיה ביולוגיים-כימיים.
- כננו-מאווררים: תעשיית הננו-אלקטרוניקה המתפתחת זקוקה נואשות לאמצעי קירור בסדרי הגודל הזעירים שבהם מדובר. בעזרת חיבור מדחף סיבובי
למנוע, אפשר למצוא פתרון לדרישה זו של התעשייה.
- כננו-משאבות: מדחף שיורכב בצורה הנכונה על גבי המנוע יוכל להדוף מולקולות. בדרך זו אפשר ליצור ננו-משאבה שתנקה תמיסות ממולקולות
בגדלים מסוימים.
- כננו-מדחפים למטרות הנעה: ננו-מדחפים כאלו יוכלו לשנע חומרים בתוך תמיסות - ואפילו להניע "צוללות" באורך של כמה מאות ננו-מטרים בזרם
הדם. מכונות עתידניות זעירות מעין אלה יוכלו לתפקד בניטור זיהומים, בשינוע חומרים ואפילו בתיקון ובשיפוץ תאים.
התוצאה הסופית היתה מעין "צוללת" שכל קוטרה ננו-מטרים אחדים. כאשר נחשפה הצוללת לאור, התמלא הכרומטופור בפרוטונים, אשר יצאו מתוכו דרך המנוע המולקולרי. מעבר הפרוטונים הביא לסיבוב של הראש ביחד עם הכנפונים הצמודים אליו, ובכך נוצר כוח הנעה. סו חיבר צוללות רבות כאלה לסיב אקטין שאורכו 10 מיקרו-מטרים, וצילם את הסיב משייט בתמיסה במהירות של עד 25 מיקרו-מטר לשנייה - מהירות גבוהה בהתחשב בסדרי הגודל שבהם מדובר.
מחקרים אלה ואחרים חשפו רק את קצה-קצהו של הפוטנציאל הגלום במנוע ה-F-ATPase Synthase. לא נותר לנו אלא להמתין ולראות, לאן ייקח אותנו העתיד ולאן יוביל הזרם - בסיועו של המדחף הזעיר, כמובן.
מנועים הנעים על מסילות: הקינזין
קיימים מנועים רבים הנעים על מסילות, וכולם חיוניים לפעילותו התקינה של התא. המורכבים מכולם הם כנראה מנועי הפולימֶראזות, הנעים על מסילת
ה-DNA כקטרים גדולים, ומשכפלים את רצף ה-DNA של התא. מנועים אלו מורכבים מתת-יחידות רבות, הקובעות היכן יתחיל מסעו של הפולימראז
והיכן יתנתק מה-DNA. הפולימראז מכיל אפילו תת-יחידה טבעתית המעגנת אותו ל-DNA, כדי לוודא שלא ירד מן המסילה בעת ביצוע תפקידו.ריבוי תת-היחידות של הפולימראז אכן מרשים, אך לרוע המזל הוא גם מקשה את חקר המנוע. לפיכך החוקרים מעדיפים להתרכז במנועים פשוטים יותר הנעים על מסילות, כגון החלבון קינֶזין.
הקינזין מורכב משני ראשים זהים, המתחברים באמצעות שני צווארים לגבעול ארוך. הקינזין מתחבר למסילה בעזרת שני הראשים שלו, ומתקדם לאורכה ב"הליכה" המזכירה את הליכתו של אדם - "רגל אחר רגל". בקצה הגבעול הקינזין נושא את המטען שלו: בועיות המכילות חומרי גלם שונים, המיועדים למקומות שונים בתא. המסילה שעליה נע הקינזין עשויה ממיקרו-טובולים - סיבים בקוטר של 24 ננו-מטר בערך. המיקרו-טובולים מתוחים בין מרכז התא לבין ממברנת התא. חלבונים ממשפחת הקינזינים והדִינֶאִינים מהלכים על המיקרו-טובולים ונושאים משאות ומטענים בכל רחבי התא.
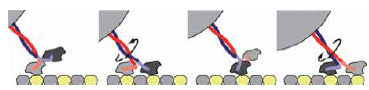
הקינזין הוא חלבון בעל שני ראשים, שבעזרתם הוא מתחבר למסילה ו"הולך" עליה
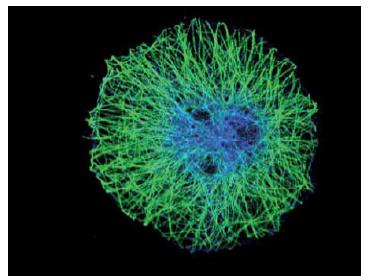
תא של דרוזופילה, שהמיקרו-טובולים מסומנים בו בירוק וה-DNA - בכחול. אפשר לראות שהמיקרו-טובולים נמתחים בין מרכז התא לבין הממברנה שלו
יתרונם הגדול של הקינזינים הוא ביציבותם. הם מסוגלים לנוע במהירות של 800 ננו-מטר לשנייה ולעשות מאות צעדים בטרם יתנתקו וייפלו מהמסילה. כל צעד שלהם מייצר 6 פיקו-ניוטון - כוח עצום ביחס לגודלם. מכיוון שכך, תפקידם בתא הוא נשיאת משאות כבדים ואברונים גדולים במיוחד.
תכונותיהם המרשימות של הקינזינים אחראיות לכך, שחוקרים רבים שואפים למצוא דרך לשלב אותם בהנדסה. את אחד המחקרים המרשימים ביותר בכיוון זה ערכו כבר ב-1999 לורן לימבריס (Limberis) וראסל סטיוארט (Stewart). צמד החוקרים טען כי אפשר לחבר כמעט 100,000 קינזינים לשטח של מיקרו-מטר רבוע אחד. מכיוון שכל מנוע ייצר כוח של כ-6 פיקו-ניוטון, סך כל הכוח שיפעילו הקינזינים יהיה בסדר גודל של עשרות ננו-ניוטון למיקרו-מטר רבוע. כוח כזה אמור להיות מסוגל להניע עצמים בסדר גודל של מיקרו-מטרים. עצמים אלה יכולים להיות גלגלי שיניים, ידיות, מיקרו-משאבות, מיקרו-גנרטורים ועוד.
כדי להוכיח את הטענה שקינזינים רבים יכולים לפעול יחד גם כשהם מסודרים בצפיפות זה לצד זה, ערכו החוקרים ניסוי מרתק במיוחד. הם הצמידו מיליוני קינזינים לשבב סיליקון ששטחו 10 מיקרו-מטרים על 10 מיקרו-מטרים, ועוביו 5 מיקרו-מטרים. אחר-כך צפו החוקרים בשבב בעזרת מיקרוסקופ אור, כאשר הוא נע על גבי מסילות מיקרו-טובולים שהוצמדו מראש לזכוכית נושאת של מיקרוסקופ. השבב נע במהירות של 800 ננו-מטר לשנייה על גבי הזכוכית, הסתובב מפעם לפעם, ואף התהפך כאשר אחת מפינותיו נתפסה במשטח. השבב שהתהפך המשיך כמובן לנוע, שכן הקינזינים היו פזורים במידה שווה גם על "גבו" של השבב...
סוג אחר של מחקר בקינזינים הוא ליצירת "פסי הנעה". כאשר מחברים את הקינזינים למשטח בעזרת קצה הגבעול, הם נותרים כששני ראשיהם פונים כלפי מעלה. כל מיקרו-טובול שיונח על גבי מסילה שכזו ינוע קדימה באמצעות הראשים הנעים של הקינזינים, בדומה לכוכב רוק הקופץ מהבמה ונע בקהל על כתפיהם וזרועותיהם של מעריציו. במבט מלמעלה נראה את המיקרו-טובול נע כנחש על גבי המסילה, אך למעשה המסילה היא המפעילה את הכוח המניע, באמצעות קינזינים רבים הדוחפים את המיקרו-טובול קדימה. מסתבר כי למסילה כזו כבר הומצאו שימושים רבים.
- פתיחת סליל DNA: ב-2003 חיבר סטפן דיאז (Diez) קצה אחד של גדיל DNA למיקרו-טובול, ואת קצהו האחר - למיקרו-טובול אחר. את שני
המיקרו-טובולים הניח על מסילות קינזינים, כך שהם נמשכו לשני כיוונים מנוגדים - וגדיל ה-DNA נפרם ונפתח.
- מדידת הכוח המופעל בין קולטנים והמולקולות המחוברות אליהם: ב-2002 חיבר הנרי הס (Hess) קולטן (רצפטור) למיקרו-טובול אחד, ואת מולקולת
היעד של אותו קולטן - למיקרו-טובול אחר שהיה מקובע בקצהו, ושימש כתמוכה בעלת קשיחות ידועה. הוא הניח לשני המיקרו-טובולים לגעת זה
בזה, כך שמולקולת היעד יכלה להתחבר לקולטן, ואז ניווט את המיקרו-טובול עם הקולטן כך שיתרחק מהתמוכה. כדי להישאר קשורה למיקרו-טובול
עם הקולטן, התמוכה נאלצה להתכופף עד שהכוחות היו גדולים מדי והקולטן התנתק מהמולקולה שהיתה קשורה לתמוכה. על-ידי צילום האירוע בזמן
אמת, הצליח הס לחשב מה מידת הכוח המחבר את הקולטן למולקולת היעד.
- מיון של מולקולות בודדות: ב-2006 יצר מרטין ג.ל. ואן דן איובל (Van Den Huevel) מסילת קינזינים שמשני צדיה היה אפשר להפעיל מתח
חשמלי. כאשר הופעל המתח, הוא השפיע על קצות המיקרו-טובולים, וגרם להם לסטות לכיוון שממנו הופעל המתח. ואן דן איובל פיזר
מיקרו-טובולים שסומנו בצבעים שונים על גבי משטח קינזינים, ובעזרת מיקרוסקופ פלואורסצנטי הצליח להפעיל את הזרם בצורה סלקטיבית, לגרום
למיקרו-טובולים השונים לנוע בכיוונים שונים ולמיין אותם לפי צבעיהם. אפשר ליישם שיטה זו גם למיון מולקולות קטנות יותר
ממיקרו-טובולים, בתנאי שהן יושבות על גבי המיקרו-טובול.
העתיד
אפשר לחשוב על המחקרים ההנדסיים שהוזכרו, ועל דומיהם, כעל המטוס הראשון של האחים רייט - לא יותר מאופניים בעלי כנפי עץ מנוסרות בגסות.
אין ספק שגם כאן לפנינו אבני דרך חשובות באמצעי ההנעה - אך בסדרי הגודל הננו-מטריים. בתוך פחות ממאה שנה, המצאתם ה"פרימיטיבית" של
האחים רייט השתכללה עד לבואינג 747 העצום, המסוגל לשאת מאות נוסעים במהירויות שהממציאים המקוריים אף לא חלמו עליהן. רק עם הזמן נדע
לאן יישאו אותנו - ואת האנושות כולה - המנועים המולקולריים-ביולוגיים למיניהם. לקריאה נוספת:
נוסח הרצאה על ATP synthase מתוך אתר אוניברסיטת אילינוי באורבנה-שמפיין
מתוך אתר פרס נובל: על פרס נובל בכימיה לשנת 1997 , שהוענק בחציו לפול ד. בויר ולג'ון א. ווקר על פענוח המנגנון האנזימי לסינתזת ATP
The ATP Synthase — A Splendid Molecular Machine, Paul D. Boyer, Annual Review of Biochemistry vol. 66:717-749
How Molecular Motors Work, James A. Spudich, Nature 372, 515-518
Molecular motors: from one motor many tails to one motor many tales. Lawrence S.B. Goldstein, Trends in Cell Biology, Vol 11, Issue 12, 477 477-482
How Molecular Motors Work, James A. Spudich, Nature 372, 515-518
Molecular motors: from one motor many tails to one motor many tales. Lawrence S.B. Goldstein, Trends in Cell Biology, Vol 11, Issue 12, 477 477-482